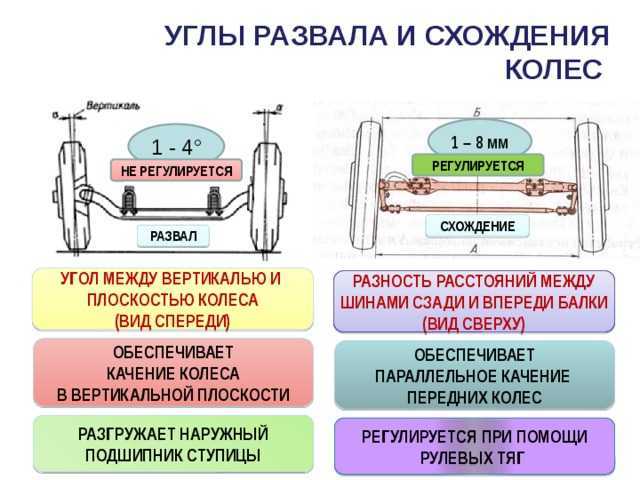
Сход развал самостоятельно не хуже стенда.
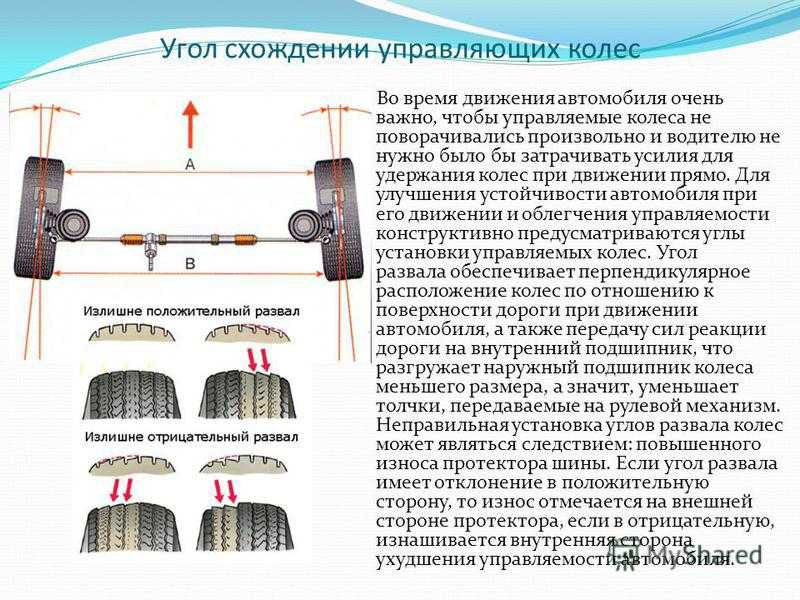
Правильно выставленные углы развала и схождения колес – одно из ключевых условий нормальной работы подвески автомобиля. Если эти параметры отклоняются от нормы, возникает ряд проблем: происходит неравномерный износ покрышек, ухудшается управляемость машины и ее курсовая устойчивость, руль смещается с нулевого положения при езде по прямой траектории. Также возрастает нагрузка на штоки амортизаторов, сайлентблоки, шрусы, элементы системы рулевого управления.
Измерение углов схода и развала должно выполняться с определенной периодичностью, в среднем 1-2 раза в год. Проверка положения колес в специализированных автосервисах — недешевая услуга, особенно при использовании компьютерных методов. Частые визиты в мастерские бьют по карману, поэтому многие автовладельцы ищут способы проведения таких работ в гаражных условиях. Предлагаем ознакомиться с простым, но точным и эффективным методом самостоятельного определения параметров схода и развала.
1 Преимущества способа
2 Важные уточнения по процедуре измерения углов установки колес
3 Три варианта построения продольной оси автомобиля
4 Измерение индивидуального схождения колес
5 Измерение углов развала
6 Измерение продольного и поперечного угла наклона оси поворота
7 Заключение
Преимущества способа
Технология, рассмотренная ниже, широко используется как начинающими, так и опытными автомобилистами. Среди ее основных преимуществ стоит выделить такие:
- Высокая точность. По этому критерию данный способ не уступает компьютерному методу. Технология позволяет измерять практически все угловые и линейные показатели, которые определяются с помощью электронной аппаратуры. Может быть учтен даже уклон площадки, на которой располагается машина во время измерения схода и развала.
- Учитываются параметры кузова. При компьютерной диагностике эти характеристики не определяются оборудованием, в лучшем случае компьютерные стенды могут вычислять величину клиренса.
 Метод, рассмотренный в статье, позволяет увидеть естественную ось симметрии кузова и определить расположение шасси относительно нее. Это поможет понять, почему после регулировок схода и развала одни колеса выступают из арок больше, чем другие.
Метод, рассмотренный в статье, позволяет увидеть естественную ось симметрии кузова и определить расположение шасси относительно нее. Это поможет понять, почему после регулировок схода и развала одни колеса выступают из арок больше, чем другие. - Вылет дисков не влияет на результаты. Параметры колесных дисков могут иметь значение при компьютерной настройке углов. Поэтому некоторые мастера, работающие в специализированных сервисах, отказываются проводить работы, если на автомобиле установлены колеса с разным вылетом на передней и задней осях. Рассматриваемый метод, в свою очередь, позволяет выполнять замеры схода и развала, даже если разные диски стоят на одной оси.
 Метод, рассмотренный в статье, позволяет увидеть естественную ось симметрии кузова и определить расположение шасси относительно нее. Это поможет понять, почему после регулировок схода и развала одни колеса выступают из арок больше, чем другие.
Метод, рассмотренный в статье, позволяет увидеть естественную ось симметрии кузова и определить расположение шасси относительно нее. Это поможет понять, почему после регулировок схода и развала одни колеса выступают из арок больше, чем другие.Важные уточнения по процедуре измерения углов установки колес
Предлагаемый метод дает возможность только определять углы установки колес (УУК), чтобы получить представление о текущих параметрах схода и развала. Дальнейшие работы по их регулировке можно проводить как своими руками (при наличии соответствующего опыта и знаний), так и в автомастерских.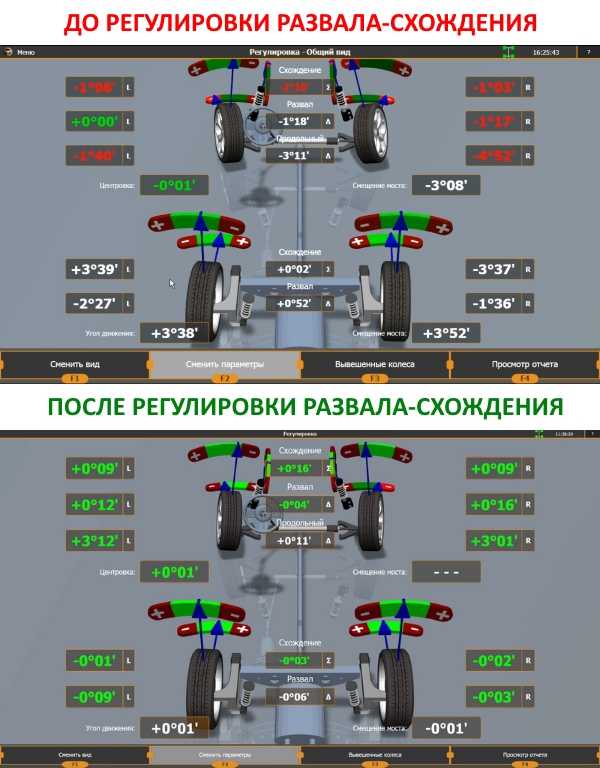
Подразумевается, что замеры проводятся при исправной подвеске и нормальном давлении в шинах. Поэтому соответствующие работы, связанные с диагностикой и ремонтом ходовой части, не рассматриваются.
Также необходимо учесть, что в данной статье не рассматривается, какими должны быть углы схода и развала. Речь пойдет только о процессе их измерений. Полученные результаты в дальнейшем нужно сравнить с заводскими значениями, которые могут отличаться для автомобилей разных марок и моделей.
Перед замерами углов схода и развала необходимо разместить автомобиль на ровной твердой поверхности – например, на бетонном полу гаража. Заезжая на рабочую площадку, следует держать руль в нулевом положении. При этом колеса будут стоять максимально близко к тому положению, в котором они находятся при движении авто по прямой дороге.
Если же автомобиль после остановки будет подниматься на домкрате или подъемнике, это приведет к разгрузке подвески, затем колеса станут на пол недостаточно ровно.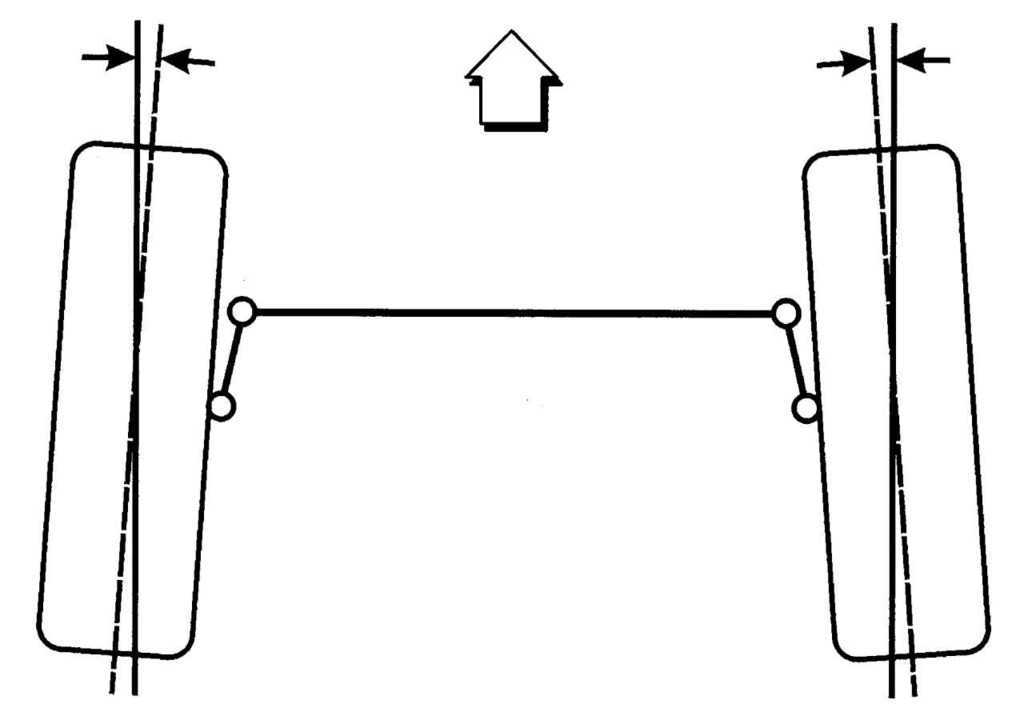 Для устранения таких погрешностей понадобятся поворотные круги или сдвижные платформы, которых большинство автовладельцев не имеют. Поэтому приподнимать кузов перед проведением замеров настоятельно не рекомендуется. Подвеска машины должна быть под привычной нагрузкой.
Для устранения таких погрешностей понадобятся поворотные круги или сдвижные платформы, которых большинство автовладельцев не имеют. Поэтому приподнимать кузов перед проведением замеров настоятельно не рекомендуется. Подвеска машины должна быть под привычной нагрузкой.
Начинать замеры следует с определения углов схода, поскольку именно эти параметры регулируются на большинстве моделей автомобилей. Данная характеристика определяется для каждого из четырех колес, поэтому ее иногда называют «полусхождением». Индивидуальный сход замеряют относительно продольной оси автомобиля. Ее можно построить несколькими способами, рассмотрим подробнее.
Три варианта построения продольной оси автомобиля
На выбор предлагаются три разных метода, с помощью которых можно построить «базовую» линию при вычислении углов индивидуального схода колес:
- В качестве продольной оси можно принять горизонтальную линию, которая проходит через ось симметрии кузова. Ее можно легко построить, отметив на переднем и заднем бамперах их средние точки.
- Второй вариант – продольная ось шасси. Эта линия выстраивается по точкам, которые располагаются в центрах расстояний между колесными ступицами.
- Довольно часто в качестве продольной оси используется линия «trast line». Она прокладывается строго в направлении движения задней оси машины. Этот вектор не всегда совпадает с продольными линиями, выстроенными по кузову или шасси, как в предыдущих вариантах. Подобное явление характерно для автомобилей, в которых задний мост немного развернут вбок. При этом машина едет боком, что часто наблюдается на старых «Жигулях», «Газелях» и др.

Угол между линией «trast line» и центральной продольной осью машины в идеальном случае должен стремиться к нулю. Но в некоторых случаях дефекты задней подвески не всегда удается устранить. Поэтому при диагностике схода и развала проводят замеры, отталкиваясь именно от «trast line».
Измерение индивидуального схождения колес
Чтобы измерить полусхождение каждого колеса, необходимо создать коридор, состоящий из двух параллельных линий. Для их создания можно использовать прочные нити, рыболовные резинки, лучи лазерных нивелиров. Линии должны проходить строго горизонтально, их нужно выстраивать по высоте центров колес. Базовый коридор можно создать тремя способами:
Для их создания можно использовать прочные нити, рыболовные резинки, лучи лазерных нивелиров. Линии должны проходить строго горизонтально, их нужно выстраивать по высоте центров колес. Базовый коридор можно создать тремя способами:
- Первый – крепление нитей непосредственно к колесам. Это самый простой, но наименее точный способ. Колеса могут иметь разный вылет относительно арок, из-за этого нарушается параллельность линий.
- Второй – фиксация нитей на стационарных опорных конструкциях. Способ более точный в сравнении с предыдущим, но неудобен тем, в процессе регулировок и прокатки автомобиля возможно смещение шасси относительно построенных линий.
- Третий – создание коридора вокруг кузова машины при помощи реечных креплений. Рейки фиксируются в передней и задней части авто, строго в центрах колес.
Есть масса примеров построения таких конструкций в англоязычном интернете. Для примера наберите в поиске «B-G rasing string lines kit».
Коридор вокруг кузова машины из речных креплений.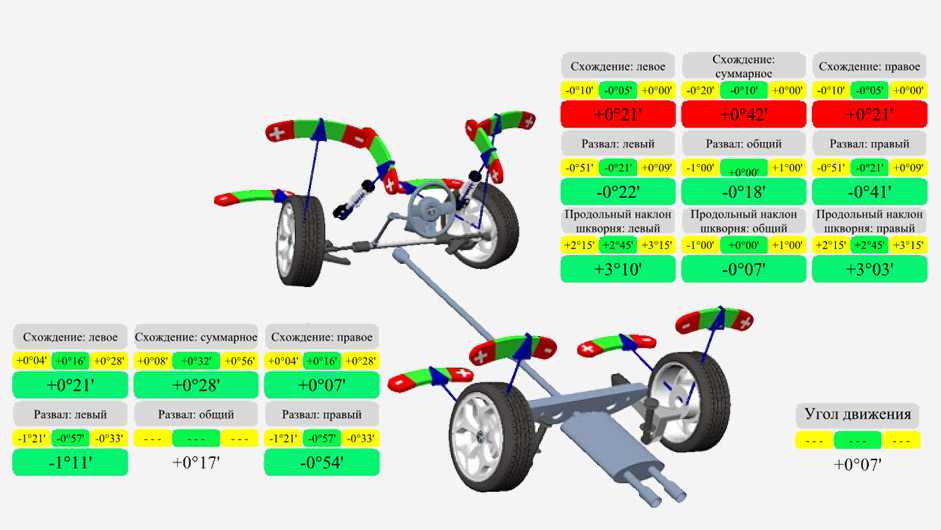
В гаражных условиях уместно использовать самый бюджетный и простой способ. Для замеров схода и развала понадобятся две алюминиевые рейки, которые можно купить в любом строительном магазине. Одну из них нужно подложить на пол у задней или передней оси авто. Далее необходимо сделать на рейке метки на небольшом расстоянии от колес — около 5 см. Затем определяется расстояние между нанесенными метками, полученное значение будет шириной измерительного коридора. Разделив этот параметр пополам, можно вычислить расположение средней линии. Далее необходимо приложить к размеченной трубке вторую рейку и перенести на нее полученные метки.
Установка меток на алюминиевых трубках.Дополнительные две метки наносятся на бампера машины ровно по центру. Их выставляют с помощью рулетки и обозначают куском изоленты или малярного скотча.
Установка метки на бампер автомобиля.Алюминиевые трубки фиксируют с помощью обычных канцелярских прищепок. Их закрепляют на бамперах, брызговиках или других элементах кузова, в зависимости от конструктивных особенностей автомобиля.
Далее необходимо натянуть на рейках в ранее отмеченных местах резиновые корды, которые можно приобрести в любом рыболовном магазине. Вместо них можно использовать неупругие нитки, которые можно закрепить на трубках резинками для денег. Это позволит избежать разрыва нитей, обеспечить необходимое натяжение и минимизировать погрешность при определении углов схода колес.
Крепление резиновых кордов к алюминиевым трубкам.На следующем этапе проводится центрирование измерительного коридора по меткам, которые были ранее обозначены на бамперах. Далее нужно выставить руль в нулевое положение, для большей точности можно использовать строительный уровень.
Установка руля в нулевое положение.После перечисленных действий уже можно обнаружить поперечное смещение колес. Для этого следует измерить расстояния от центров ступиц до нитей базового коридора и определить разницу между ними. Это актуально для водителей, в чьих автомобилях колеса выходят из арок на разные расстояния.
Теперь можно приступать к измерению индивидуального схода каждого колеса. Замеры проводятся путем определения расстояний от линий базового коридора до симметричных точек обода дисков спереди и сзади от центра колеса. Полученные результаты необходимо зафиксировать в тетради или, за неимением оной, записать мелом на асфальте в гараже.
Определение расстояний от линий базового коридора до симметричных точек обода дисков.Затем необходимо прокатать автомобиль вперед или назад на пол оборота колеса, не меняя положения линий измерительного коридора. Важно, чтобы руль при этом оставался в прежнем положении. Далее нужно несколько раз провести повторные замеры индивидуального схождения колес. Если между двумя измерениями нет существенных различий, их можно принять за истинные. Если разница значительна, следует прокатать машину еще на несколько десятков сантиметров и обнаружить «кривые» колеса. Таким образом вычисляем кривые колеса. Чтобы компенсировать биения колес при расчетах, находим наиболее часто повторяющиеся значения индивидуальных схождений.
Чтобы компенсировать биения колес при расчетах, находим наиболее часто повторяющиеся значения индивидуальных схождений.
Для пересчета миллиметров в градусы и минуты воспользуемся таблицей ниже.
Таблица перевода миллиметров индивидуального схождения колеса в градусы и минуты.Когда все углы полусхождения определены с достаточной точностью, можно вычислить угол разворота задней оси. О наличии такого угла могут свидетельствовать различия параметров полусхождения задних колес относительно центральной линии.
Если же регулировка задних колес не планируется, можно переориентировать базовый коридор по trast line, проложенной через заднюю ось. Для этого нужно сдвинуть закрепленные рейки таким образом, чтобы полусхождения задних колес выровнялись по новому положению коридора. Затем необходимо будет откорректировать по нему положение передних колес, зафиксировав руль автомобиля строго по центру. Таким образом машина будет ехать немного боком, но водитель не будет замечать этого.
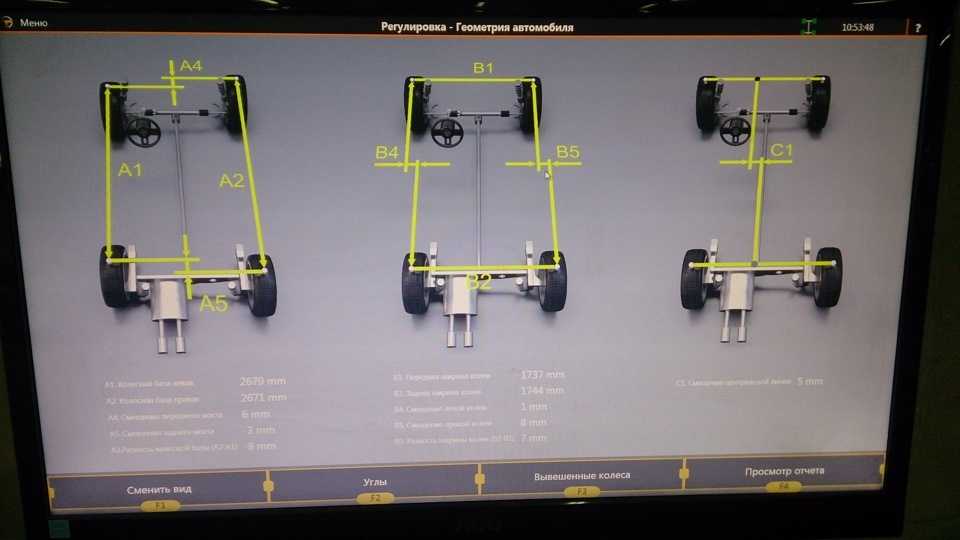
При определении параметров полусхождения необходимо учитывать суммарное схождение колёс на осях (на передней оси и на задней), которое складывается из индивидуальных углов с учетом их знаков. Именно этот показатель определяет, будет ли равномерно изнашиваться резина, и насколько управляемым станет автомобиль во время движения. Чтобы узнать оптимальное значение общего схождения для конкретной модели авто, следует изучить справочные материалы, в которых указаны рекомендации производителя.
Именно этот показатель определяет, будет ли равномерно изнашиваться резина, и насколько управляемым станет автомобиль во время движения. Чтобы узнать оптимальное значение общего схождения для конкретной модели авто, следует изучить справочные материалы, в которых указаны рекомендации производителя.
Измерение углов развала
Для определения этих параметров базовый коридор не понадобится, его можно убрать. Углы развала можно определить несколькими способами: с помощью отвеса, пузырькового уровня, цифрового инклинометра или другого угломера.
Перед началом замера углов развала нужно удостовериться, что машина стоит на ровной поверхности. Если же развал будет измеряться с помощью цифрового инклинометра, то достаточно откалибровать прибор по наклону поверхности, на которой расположена машина. Для этого следует приложить к поверхности угольник и сверить расположение угломера с положением его вертикальной грани.
На ободе колеса ставим маркером или мелом две вертикальные метки.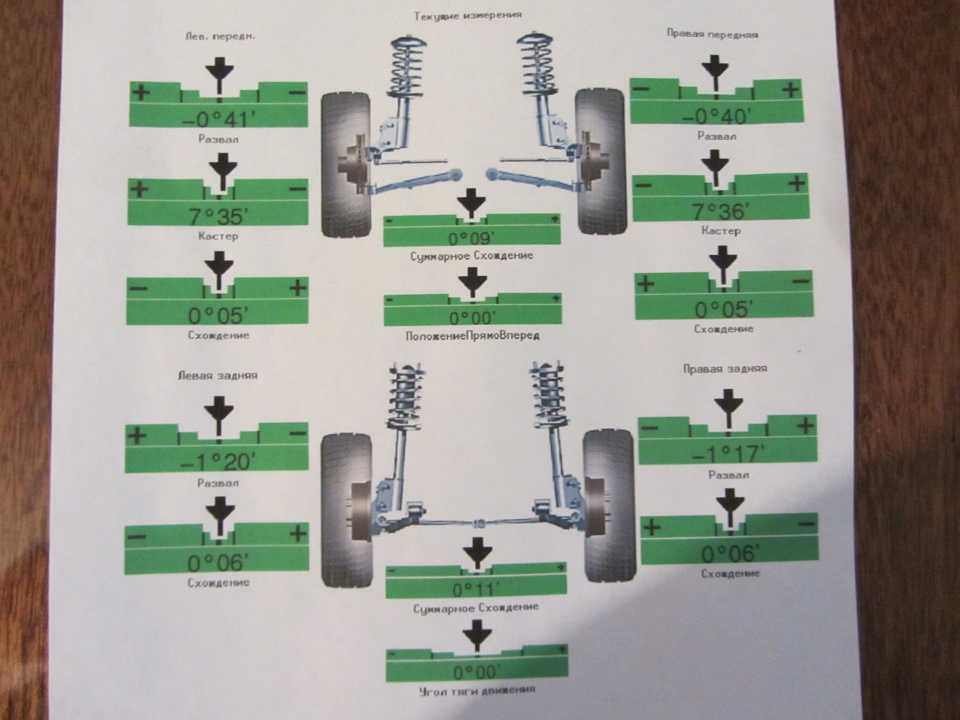 Шнур с отвесом прикладываем к крылу, и вдоль меток линейкой или штангенциркулем измеряем расстояние от обода до шнура у верхней метки, а затем у нижней. Разница должна быть в пределах +-2мм. Прокатываем машину на четверть оборота колеса (90 градусов) и делаем еще две вертикальные метки. Снова замеряем расстояние от отвеса до обода колеса, тем самым нивелируем погрешность измерения, связанную с биением диска колеса.
Шнур с отвесом прикладываем к крылу, и вдоль меток линейкой или штангенциркулем измеряем расстояние от обода до шнура у верхней метки, а затем у нижней. Разница должна быть в пределах +-2мм. Прокатываем машину на четверть оборота колеса (90 градусов) и делаем еще две вертикальные метки. Снова замеряем расстояние от отвеса до обода колеса, тем самым нивелируем погрешность измерения, связанную с биением диска колеса.
Для переднеприводных автомобилей усреднённые значения углов развала считаются в норме 0 ± 1 мм, для заднего привода нормой считается диапазон +1 ±3 мм. Для дисков размеров 13 и 14 дюймов 1 миллиметр развала равен примерно 10 угловым минутам. Точную величину допуска развала колес для конкретного автомобиля лучше всего узнать из рекомендаций завода изготовителя автомобиля.
Измерение угла развала с помощью пузырькового уровня.Угол развала также можно измерить с помощью пузырькового уровня или телефона с установленной программой для измерения углов.
Измерение продольного и поперечного угла наклона оси поворота
Наибольший интерес представляет измерение параметров наклона поворотных осей. Их положение определяется двумя ключевыми характеристиками: поперечным наклоном и продольным (кастером).
Короткое видео (2 мин) о том, что такое кастер
Если бы колеса авто могли выворачиваться на 90 градусов и становиться поперек кузова, эти параметры можно было бы легко вычислить с помощью инклинометра. Но в реальных условиях этого сделать невозможно, поэтому нужно ограничиваться поворотом на 20 градусов в обе стороны относительно нулевого положения. Разницу полученных углов нужно умножить на дополнительный коэффициент 1,5 – это необходимо для компенсации погрешностей, которые допускаются в процессе замеров.
Чтобы определить кастер, сначала нужно развернуть каждое колесо на 20 градусов перед осью поворота: правое влево, а левое вправо. При этом нужно выставить угломер на ноль или записать полученные результаты. Далее колеса разворачивают за ось поворота: правое вправо, левое влево. Если угол, измеренный позади оси поворота больше угла измеренного перед осью поворота, то кастер положительный. Если наоборот, то кастер отрицательный. Угломер при этом должен показывать небольшую разницу в несколько градусов. Эту разность следует умножить на тот же коэффициент 1,5. К примеру, если угломер показал значение +2 градуса, то величину кастера нужно принять за +3 градуса.
Аналогичным способом вычисляют поперечный наклон, но при этом угломер устанавливают в несколько ином положении. Для дополнительного удобства можно использовать смартфон, на который следует заранее установить приложение, моделирующее два взаимно перпендикулярных пузырьковых уровня.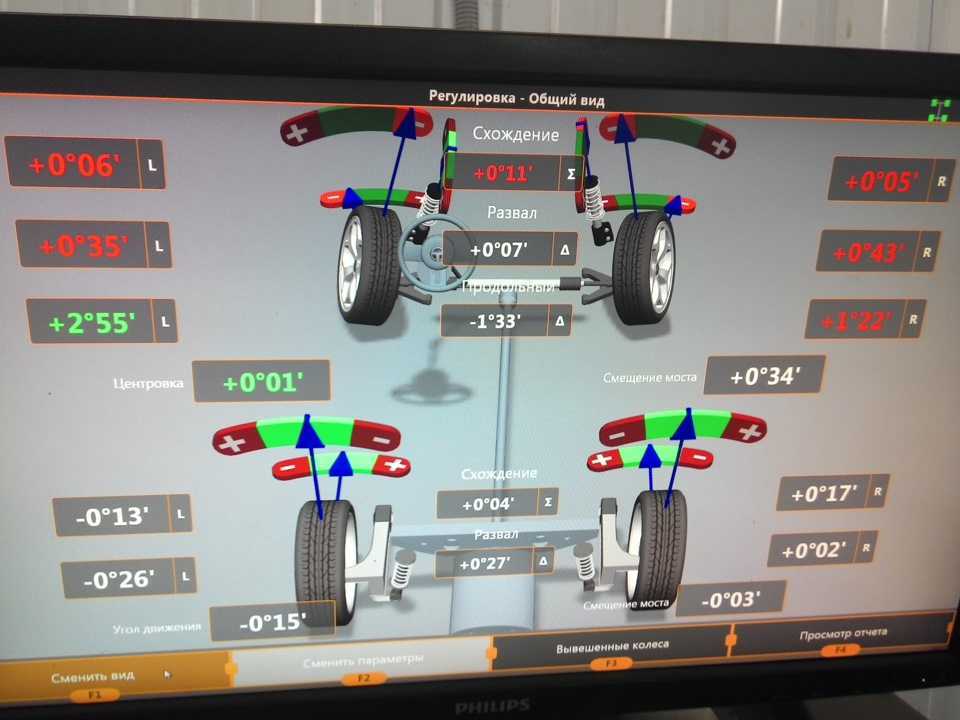 Также для этих целей сгодится и обычный строительный уровень, желательно с вращающимся пузырьком.
Также для этих целей сгодится и обычный строительный уровень, желательно с вращающимся пузырьком.
Если задача заключается не в измерении конкретных числовых значений наклонов, а лишь в их сравнении (они всё равно не регулируются на большинстве автомобилей), то результаты не обязательно умножать на поправочный коэффициент 1,5. Главное – убедиться в том, что углы равны на обеих полуосях (справа и слева).
В процессе самостоятельных замеров не обязательно строго соблюдать поворот колес ровно на 20 градусов, особенно когда нет соответствующих инструментов для определения этих углов (например, проградуированных поворотных кругов). Можно выворачивать руль примерно на 3/4 оборота. Необходимо следить, чтобы при повороте влево и вправо руль фиксировался в симметричных положениях. Для этого можно нанести какие-либо пометки в нижней части рулевого колеса.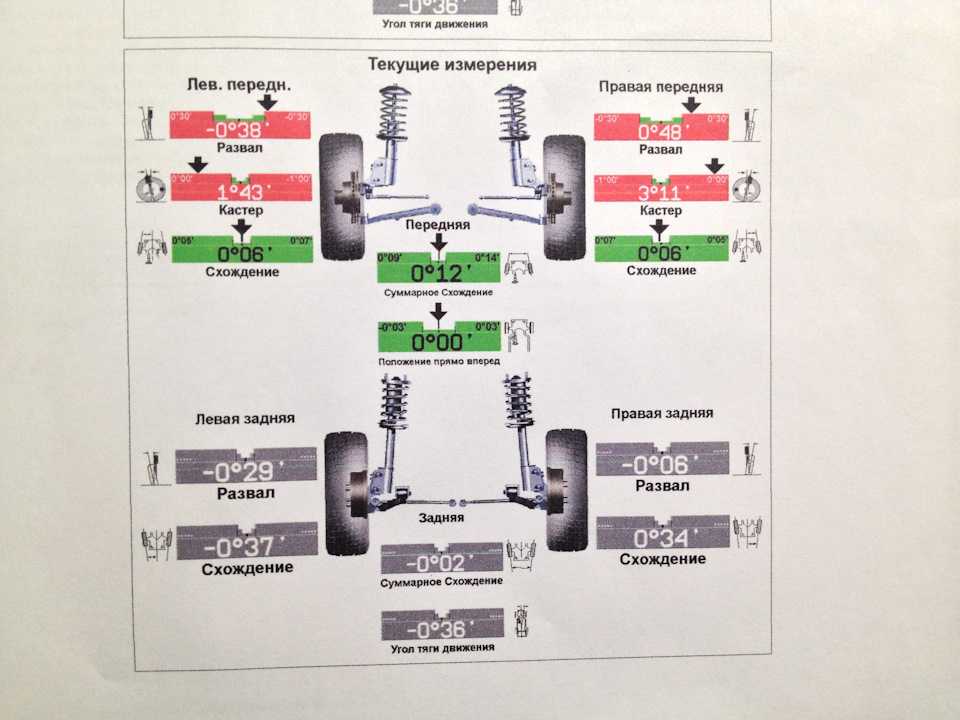 При нулевом положении руля линия, соединяющая метки, должна проходить строго горизонтально.
При нулевом положении руля линия, соединяющая метки, должна проходить строго горизонтально.
Еще один способ добиться точных результатов – начертить углы на асфальте возле колес при помощи транспортира. При отсутствии этого инструмента можно использовать квадратный лист бумаги с равными сторонами. Сложив его по диагонали, получим угол 45 градусов. Затем один угол снова складывается пополам, получается угол около 22,5 градуса, что близко к стандартным 20. Далее лист нужно приложить к колесу и очертить его грань мелом, тем самым создав нужные пометки на асфальте.
Разметка углов для поворота колесаЗаключение
Способы измерения УУК автомобиля, рассмотренные в данной статье, практически не используются на профессиональных автосервисах. Компьютерный метод более удобен и эффективен, требует меньших затрат времени. Но в быту автомобилистов часто бывают ситуации, когда визит на СТО затруднен по разным причинам: например, если машина временно не на ходу, не хватает финансов, или когда водителю нужно лишь убедиться в нормальном состоянии подвески машины.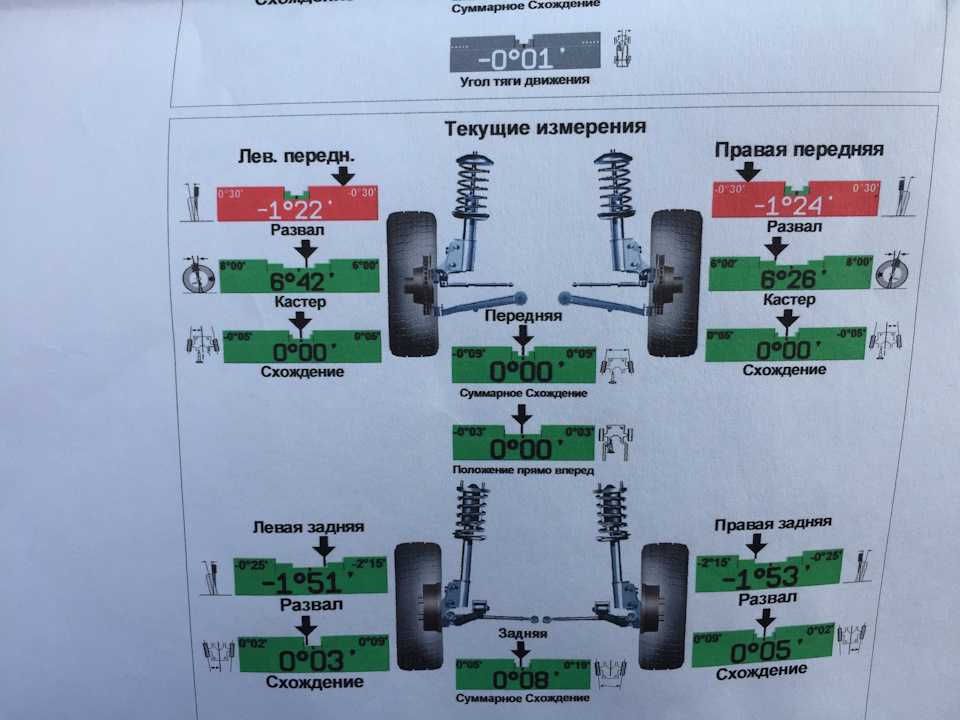 В таких ситуациях предложенная технология оптимально подойдет для замера углов схода/развала и станет достойной альтернативой компьютерной диагностике УУК.
В таких ситуациях предложенная технология оптимально подойдет для замера углов схода/развала и станет достойной альтернативой компьютерной диагностике УУК.
В заключении статьи рекомендую посмотреть видео.
Развал схождение своими руками | Премиум Газ Сервис
При отсутствии времени или денег на поездку в СТО сход развал сделать самому тоже возможно, но для этого понадобится полный комплект инструментов. Кроме того, сперва нужно проверить ходовую, чем и требуется заняться в первую очередь.
Необходимо проверить все составляющие узлов ходовой, если хоть какая-то деталь требует замены или ремонта, регулировку положения колес пока делать бессмысленно. Только после всех ремонтных работ стоит заняться самостоятельной настройкой развал схождения.
Перед проверкой также стоит оценить степень накаченности шин и загрузки автомобиля, что также может повлиять на измерения перед изменением положения колес. Теперь более подробно о том, как самому сделать развал схождение в гаражных условиях.
Инструментарий
Чтобы выполнить все необходимые манипуляции понадобится следующий набор:
- Смотровая яма, самодельная платформа или подъемник.
- Мелок для меток.
- Домкрат.
- Линейка, рулетка, (отлично подойдет строительный уровень) нить.
- Гаечные ключи для разбора узла колеса и части рулевой стойки.
- Техпаспорт авто.
Это минимальный набор, который необходим чтобы сделать сход развал самому. Машина обязательно должна находится на ровной поверхности, чтобы измерения были максимально точными. Чтобы начать работу необходимо зафиксировать свое транспортное средство так, чтобы колеса заняли положение, как при поездке с линейной прямой дорогой. Рекомендуется использовать направляющие в качестве ориентира, например длинные прямые бруски или уголки. Параметры развал схождения определяет стенд с компьютерной аппаратурой при помощи датчиков, установленных на колесах, а саму подгонку выполняют мастера. Все замеры чрезвычайно точные, благодаря чему можно определить даже незначительный дефект в положении колеса. Чтобы данные были наиболее достоверными перед началом процедуры проверяют состояние ходовой, давление в шинах и наличие груза в авто.
Параметры развал схождения определяет стенд с компьютерной аппаратурой при помощи датчиков, установленных на колесах, а саму подгонку выполняют мастера. Все замеры чрезвычайно точные, благодаря чему можно определить даже незначительный дефект в положении колеса. Чтобы данные были наиболее достоверными перед началом процедуры проверяют состояние ходовой, давление в шинах и наличие груза в авто.
Поэтапное проведение корректировки колес
Теперь, как сделать развал схождение своими руками. Короткая инструкция для корректировки развала.
- К крылу авто потребуется прикрепить нить с грузом чтобы проверить перпендикулярность колеса. Нить протягивается через центр колеса и крепится на другой стороне. Снизу шины делается первая пометка, сверху через пол оборота вторая.
- Линейкой или рулеткой измеряем расстояние от нити до обоих пометок, данные фиксируем удобным способом. Интересует нас разница между верхним и нижним показателем. В техпаспорте прописана норма, сравниваем. Если цифры совпадают, регулировка не нужна.
- Процедуру сверки нужно провести для двух пар колес или только для пары управления.
- При обнаружении несоответствий потребуется отодвинуть поворотный кулачок колеса, а для этого придется снять колесо. Подгоняем кулачок в нужное положение ослабив его крепежные гайки. Может понадобится несколько попыток.
 Если цифры совпадают, регулировка не нужна.
Если цифры совпадают, регулировка не нужна.Для регулировки схождения нужно провести четыре измерения:
- Два замера внутреннего расстояния колес между их самыми близкими и самыми дальними точками.
- Еще два внешних расстояния, от крайних точек к направляющим.
- Проверить показатели и сравнить их с данными в паспорте.
- При наличии отклонений от нормы — расхождения в замерах между точками в пункте 1 и 2 составляет более 10%, то требуется подгонка параметров, выполняется она вручную.
Чтобы подкорректировать сход колес требуется удлинять или укорачивать рулевые тяги. Для этого придется предпринять несколько попыток, после чего придется еще раз провести контрольные замеры.
Лучше обращайтесь к специалистам
Во всяком случае, развал сход своими руками делать строго не рекомендуется из-за большого риска неверной настройки. Проще все-таки выделить немного времени и посетить ближайшее СТО для того, чтобы мастера при помощи современной точной аппаратуры провели диагностику. Кроме того, для техобслуживания применяется специальный инструмент, который значительно упрощает и ускоряет работу, поэтому получится еще немного сэкономить времени, хоть и придется потратить немного денег.
GAN — Способы повышения производительности GAN | by Jonathan Hui
Фото Andy BealesМодели GAN могут сильно пострадать в следующих областях по сравнению с другими глубокими сетями.
- Несходимость : модели не сходятся и, что еще хуже, становятся нестабильными.
- Сбой режима : генератор выдает ограниченные режимы, и
- Медленное обучение: исчез градиент для обучения генератора.

В этой статье, посвященной GAN, рассматриваются способы улучшения GAN. В частности,
- Измените функцию стоимости для лучшей цели оптимизации.
- Добавьте дополнительные штрафы к функции стоимости для обеспечения соблюдения ограничений.
- Избегайте самоуверенности и переобучения.
- Улучшенные способы оптимизации модели.
- Добавить метки.
Но имейте в виду, что это динамичная тема, так как исследования продолжаются очень активно.
Генератор пытается найти лучшее изображение, чтобы обмануть дискриминатор. «Лучший» образ продолжает меняться, когда обе сети противодействуют своему оппоненту. Однако оптимизация может оказаться слишком жадной и превратить ее в бесконечную игру в кошки-мышки. Это один из сценариев, когда модель не сходится и мода рушится.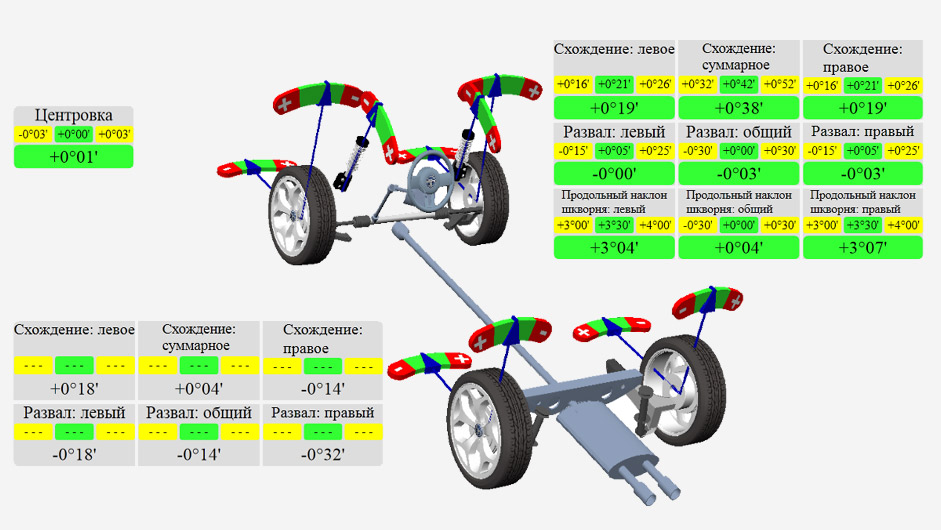
Сопоставление признаков изменяет функцию стоимости для генератора, минимизируя статистическую разницу между признаками реальных изображений и сгенерированных изображений. Часто мы измеряем L2-расстояние между средними значениями их векторов признаков. Таким образом, сопоставление признаков расширяет цель от победы над противником до сопоставления признаков на реальных изображениях. Вот новая целевая функция:
, где f(x) — это вектор признаков, извлеченный в непосредственном слое дискриминатором.
Средние значения характеристик реального изображения вычисляются для каждой мини-партии, которые колеблются в каждой партии. Это хорошая новость в смягчении коллапса режима. Это вводит случайность, из-за которой дискриминатору труднее переобучить себя.
Сопоставление функций эффективно, когда модель GAN нестабильна во время обучения.
При сворачивании режима все созданные изображения выглядят одинаково. Чтобы смягчить проблему, мы загружаем в дискриминатор реальные изображения и сгенерированные изображения отдельно в разных пакетах и вычисляем сходство изображения x с изображениями в одном пакете. Мы добавляем сходство o(x) в один из плотных слоев дискриминатора, чтобы классифицировать, является ли это изображение реальным или сгенерированным.
Чтобы смягчить проблему, мы загружаем в дискриминатор реальные изображения и сгенерированные изображения отдельно в разных пакетах и вычисляем сходство изображения x с изображениями в одном пакете. Мы добавляем сходство o(x) в один из плотных слоев дискриминатора, чтобы классифицировать, является ли это изображение реальным или сгенерированным.
Если мод начинает схлопываться, сходство сгенерированных изображений увеличивается. Дискриминатор может использовать эту оценку для обнаружения сгенерированных изображений и наказания генератора, если режим рушится.
Сходство o(xi) между изображением xi и другие изображения в том же пакете вычисляются с помощью матрицы преобразования T . Уравнения немного сложно отследить, но концепция довольно проста. Но не стесняйтесь переходить к следующему разделу, если хотите.
На рисунке выше xi — входное изображение, а xj — остальные изображения в том же пакете.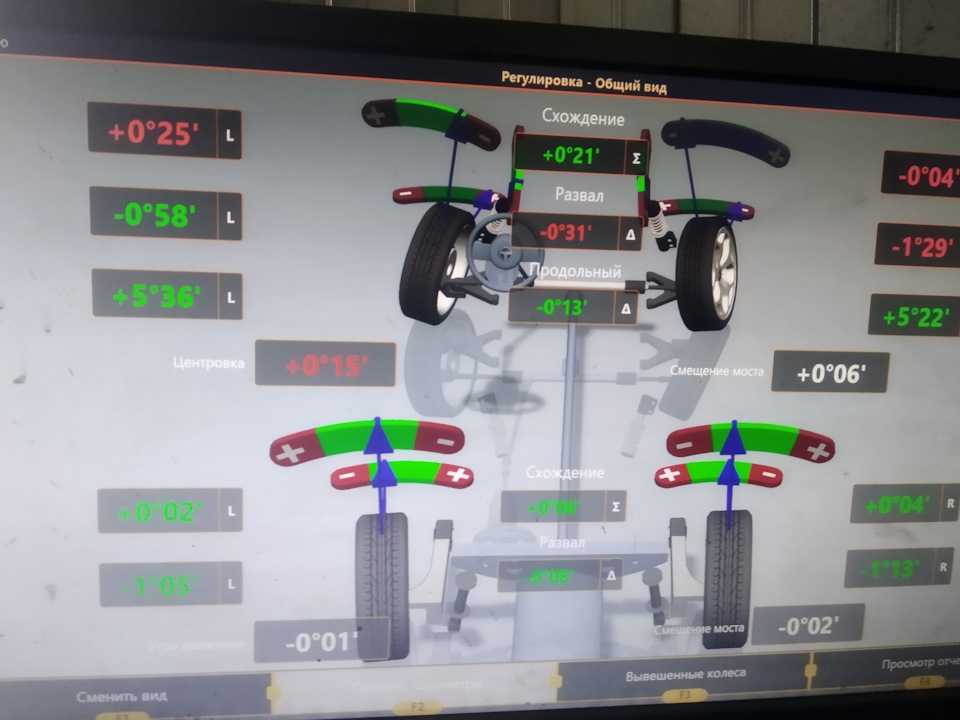 Мы используем матрицу преобразования T для преобразования функций 9от 0040 xi до Mi , который представляет собой матрицу B×C.
Мы используем матрицу преобразования T для преобразования функций 9от 0040 xi до Mi , который представляет собой матрицу B×C.
Мы получаем сходство c(xi, xj) между изображением i и j , используя L1-норму и следующее уравнение.
Сходство o(xi) между изображением xi и остальными изображениями в пакете составляет
Дискриминация в мини-пакетах позволяет нам очень быстро создавать визуально привлекательные образцы, и в этом отношении она превосходит сопоставление признаков.
Глубинные сети могут страдать от самоуверенности. Например, он использует очень мало признаков для классификации объекта. Чтобы смягчить проблему, глубокое обучение использует регулирование и отсев, чтобы избежать чрезмерной уверенности.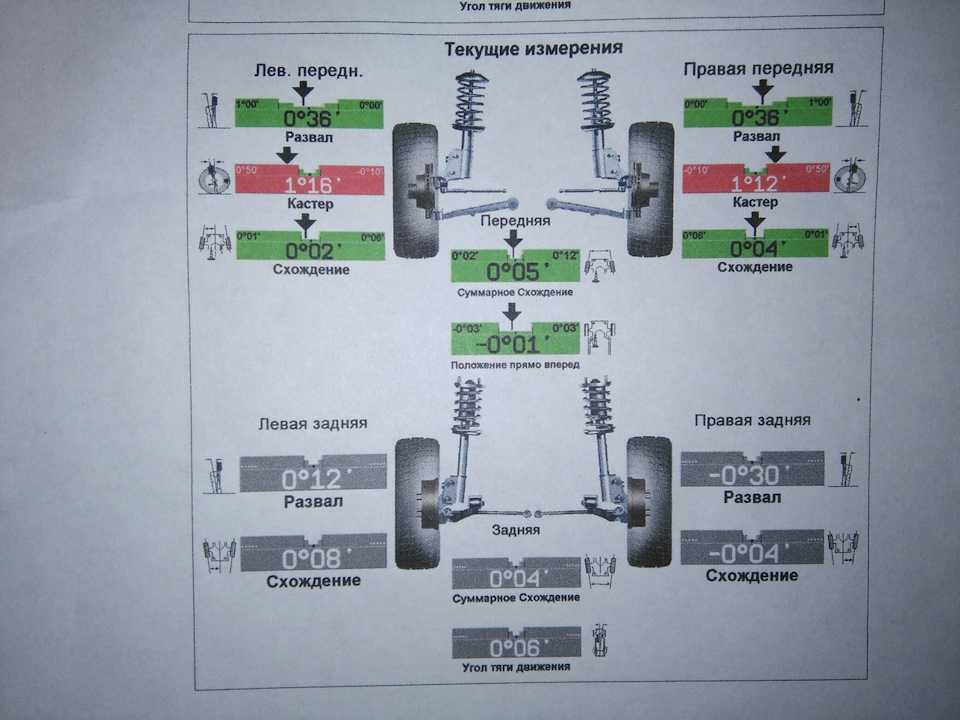
В GAN, если дискриминатор зависит от небольшого набора функций для обнаружения реальных изображений, генератор может просто создавать эти функции только для использования дискриминатора. Оптимизация может оказаться слишком жадной и не даст долгосрочной выгоды. В GAN излишняя самоуверенность очень вредит. Чтобы избежать этой проблемы, мы штрафуем дискриминатор, когда предсказание для любых реальных изображений превышает 0,9.( D (реальное изображение) > 0,9 ). Это делается путем установки нашего целевого значения метки равным 0,9 вместо 1,0. Вот псевдокод:
p = tf.placeholder(tf.float32, shape=[None, 10])# Используйте 0,9 вместо 1,0.
feed_dict = {
p: [[0, 0, 0, 0.9, 0, 0, 0, 0, 0, 0]] # Изображение с меткой "3"
}# logits_real_image - это логиты, вычисленные
# дискриминатором для реальных изображений.
d_real_loss = tf.nn.sigmoid_cross_entropy_with_logits(
labels=p, logits=logits_real_image)
При историческом усреднении мы отслеживаем параметры модели для последних моделей t .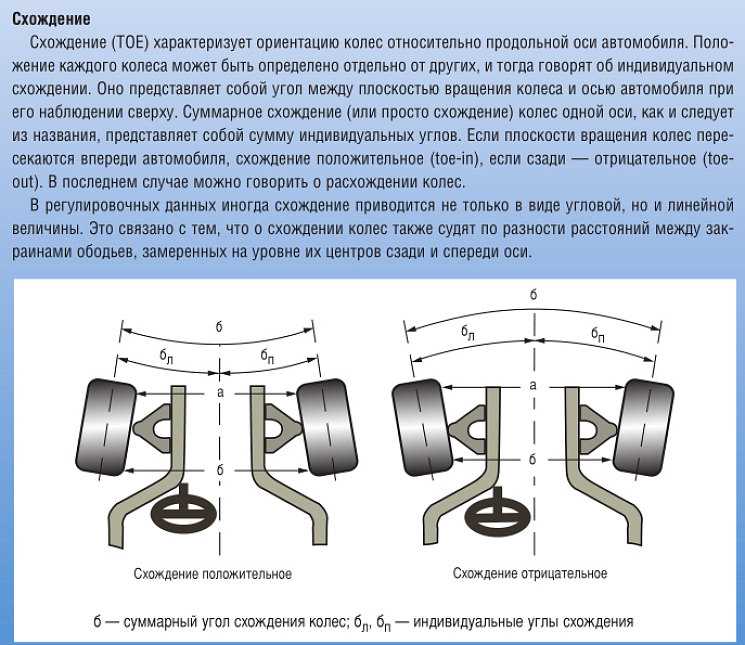 В качестве альтернативы мы обновляем скользящее среднее параметров модели, если нам нужно сохранить длинную последовательность моделей.
В качестве альтернативы мы обновляем скользящее среднее параметров модели, если нам нужно сохранить длинную последовательность моделей.
Мы добавляем приведенную ниже стоимость L2 к функции стоимости, чтобы оштрафовать модель, отличающуюся от среднего исторического значения.
Для GAN с невыпуклой функцией объекта историческое усреднение может остановить круг моделей вокруг точки равновесия и действовать как демпфирующая сила для сходимости модели.
Оптимизация модели может быть слишком жадной для подавления того, что в данный момент генерирует генератор. Чтобы решить эту проблему, воспроизведение опыта поддерживает самые последние сгенерированные изображения из прошлых итераций оптимизации. Вместо подбора моделей только текущими сгенерированными изображениями мы также подаем на дискриминатор все последние сгенерированные изображения. Следовательно, дискриминатор не будет переобучен для конкретного временного экземпляра генератора.
Многие наборы данных снабжены метками для типов объектов их выборок. Тренировать ГАН уже тяжело. Таким образом, любая дополнительная помощь в обучении GAN может значительно улучшить производительность. Добавление метки как части скрытого пространства z помогает обучению GAN. Ниже показан поток данных, используемый в CGAN для использования меток в образцах.
Тренировать ГАН уже тяжело. Таким образом, любая дополнительная помощь в обучении GAN может значительно улучшить производительность. Добавление метки как части скрытого пространства z помогает обучению GAN. Ниже показан поток данных, используемый в CGAN для использования меток в образцах.
Имеют ли значение функции затрат? Так и должно быть, иначе все эти исследовательские усилия окажутся напрасными. Но если вы услышите о статье Google Brain за 2017 год, у вас обязательно возникнут сомнения. Но повышение качества изображения по-прежнему остается главным приоритетом. Скорее всего, мы увидим, как исследователи пробуют разные функции затрат, прежде чем мы получим точный ответ на вопрос о достоинствах.
На следующем рисунке перечислены функции стоимости для некоторых распространенных моделей GAN.
Таблица изменена отсюда. Мы решили не детализировать эти функции стоимости в этой статье. Вот статьи, в которых подробно рассматриваются некоторые общие функции затрат: WGAN/WGAN-GP, EBGAN/BEGAN, LSGAN, RGAN и RaGAN. В конце этой статьи мы перечисляем статью, в которой все эти функции стоимости изучаются более подробно. Поскольку функция стоимости является одной из основных областей исследований в GAN, мы рекомендуем вам прочитать эту статью позже.
В конце этой статьи мы перечисляем статью, в которой все эти функции стоимости изучаются более подробно. Поскольку функция стоимости является одной из основных областей исследований в GAN, мы рекомендуем вам прочитать эту статью позже.
Вот некоторые оценки FID (чем ниже, тем лучше) для некоторых наборов данных. Это один ориентир, но имейте в виду, что еще слишком рано делать какие-либо выводы о том, какие функции затрат работают лучше всего. Действительно, пока не существует единой функции стоимости, которая бы работала лучше всего среди всех различных наборов данных.
Источник(MM GAN — это функция стоимости GAN в оригинальной статье. NS GAN — это альтернативные функции стоимости, учитывающие исчезающие градиенты в той же статье.)
Но ни одна модель не работает хорошо без хороших гиперпараметров, а настройка GAN требует времени. Будьте терпеливы в оптимизации гиперпараметров, прежде чем случайным образом тестировать различные функции стоимости. Некоторые исследователи предположили, что настройка гиперпараметров может принести большую отдачу, чем изменение функций затрат.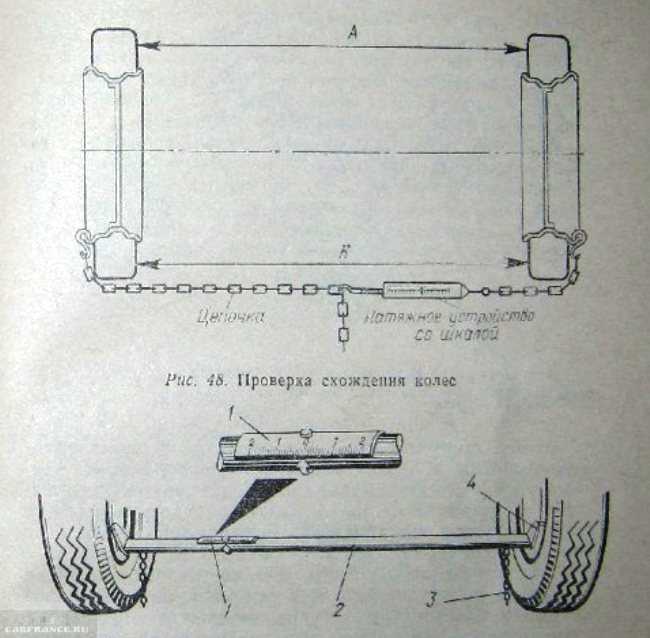 Тщательно настроенная скорость обучения может смягчить некоторые серьезные проблемы GAN, такие как сбой режима. В частности, снизьте скорость обучения и повторите обучение, когда произойдет сбой режима.
Тщательно настроенная скорость обучения может смягчить некоторые серьезные проблемы GAN, такие как сбой режима. В частности, снизьте скорость обучения и повторите обучение, когда произойдет сбой режима.
Мы также можем поэкспериментировать с разными скоростями обучения для генератора и дискриминатора. Например, на следующем графике используется скорость обучения 0,0003 для дискриминатора и 0,0001 для генератора в обучении WGAN-GP.
Источник- Масштабируйте значение пикселя изображения от -1 до 1. Используйте tanh в качестве выходного слоя для генератора.
- Экспериментальная выборка z с распределением Гаусса.
- Пакетная нормализация часто стабилизирует обучение.
- Использовать PixelShuffle и транспонировать свертки для повышения дискретизации.
- Избегайте максимального объединения для понижения частоты дискретизации. Используйте сверточный шаг.
- Оптимизатор Adam обычно работает лучше других методов.
- Добавьте шум к реальным и сгенерированным изображениям перед их подачей в дискриминатор.
Динамика моделей GAN еще недостаточно изучена. Таким образом, некоторые советы являются всего лишь предложениями, и пробег может варьироваться. Например, в документе LSGAN сообщается, что RMSProp имеет более стабильное обучение в своих экспериментах. Это довольно редкое явление, но оно демонстрирует трудности с составлением общих рекомендаций.
Дискриминатор и генератор постоянно конкурируют друг с другом. Будьте готовы к тому, что значение функции стоимости может увеличиваться и уменьшаться. Не прекращайте обучение преждевременно, даже если может показаться, что стоимость растет. Визуально контролируйте результаты, чтобы проверить ход обучения.
Пакетная нормализация BM становится стандартом де-факто во многих проектах глубоких сетей. Среднее значение и дисперсия BM получены из текущей мини-партии. Однако это создает зависимость между образцами. Сгенерированные изображения не являются независимыми друг от друга.
Это отражено в экспериментах, в которых сгенерированные изображения демонстрируют цветовой оттенок в одной и той же партии.
Оранжевый оттенок на верхней партии и зеленоватый оттенок на второй. ИсточникПервоначально мы выбираем z из случайного распределения, которое дает нам независимые выборки. Однако смещение, создаваемое пакетной нормализацией, подавляет случайность z .
Нормализация виртуальной партии (VBN) отбирает контрольную партию перед обучением. На прямом проходе мы можем предварительно выбрать эталонную партию для вычисления параметров нормализации ( μ и σ) для BN. Однако мы подгоним модель к этой эталонной партии, поскольку используем одну и ту же партию для всего обучения. Чтобы смягчить это, мы можем объединить эталонную партию с текущей партией для вычисления параметров нормализации.
Случайные начальные значения, используемые для инициализации параметров модели, влияют на производительность GAN. Как показано ниже, оценки FID при измерении производительности GAN различаются в 50 отдельных прогонах (обучение). Но диапазон относительно невелик и, вероятно, будет выполнен только в более поздней точной настройке.
Как показано ниже, оценки FID при измерении производительности GAN различаются в 50 отдельных прогонах (обучение). Но диапазон относительно невелик и, вероятно, будет выполнен только в более поздней точной настройке.
В документе Google Brain указано, что LSGAN иногда дает сбой или рушится в некоторых наборах данных, и необходимо перезапустить обучение с другим случайным начальным числом.
DGCAN настоятельно рекомендует добавить BM в проект сети. Использование BM также стало общей практикой во многих моделях глубоких сетей. Однако будут исключения. На следующем рисунке показано влияние BN на различные наборы данных. Ось y представляет собой оценку FID, которая чем ниже, тем лучше. Как указано в документе WGAN-GP, при использовании BN должен быть выключен. Мы предлагаем читателям проверить используемую функцию стоимости и соответствующую производительность FID на BN, а также проверить настройку с помощью экспериментов.
Изменено из исходного кода. Спектральная нормализация — это нормализация веса, которая стабилизирует обучение дискриминатора. Он управляет константой Липшица дискриминатора, чтобы смягчить проблему взрывающегося градиента и проблему коллапса моды. Концепция в значительной степени основана на математике, но концептуально она ограничивает изменения веса на каждой итерации, а не зависит от небольшого набора признаков при различении изображений дискриминатором. Этот подход будет легким в вычислительном отношении по сравнению с WGAN-GP и обеспечит хорошее покрытие режима, которое преследует многие методы GAN.
Он управляет константой Липшица дискриминатора, чтобы смягчить проблему взрывающегося градиента и проблему коллапса моды. Концепция в значительной степени основана на математике, но концептуально она ограничивает изменения веса на каждой итерации, а не зависит от небольшого набора признаков при различении изображений дискриминатором. Этот подход будет легким в вычислительном отношении по сравнению с WGAN-GP и обеспечит хорошее покрытие режима, которое преследует многие методы GAN.
Крах режима может быть не так уж и плох. Качество изображения часто улучшается, когда режим рушится. Фактически, мы можем собрать лучшую модель для каждого режима и использовать их для воссоздания различных режимов изображений.
Источник Дискриминатор и генератор всегда находятся в перетягивании каната, чтобы подорвать друг друга. Коллапс моды и уменьшение градиента часто объясняют дисбалансом между дискриминатором и генератором. Мы можем улучшить GAN, обратив внимание на балансировку потерь между генератором и дискриминатором. К сожалению, решение кажется неуловимым. Мы можем поддерживать статическое соотношение между количеством итераций градиентного спуска на дискриминаторе и генераторе. Даже это кажется привлекательным, но многие сомневаются в его пользе. Часто мы поддерживаем соотношение один к одному. Но некоторые исследователи также проверяют соотношение 5 итераций дискриминатора на одно обновление генератора. Также предлагается сбалансировать обе сети с помощью динамической механики. Но только в последние годы мы получили некоторую поддержку.
К сожалению, решение кажется неуловимым. Мы можем поддерживать статическое соотношение между количеством итераций градиентного спуска на дискриминаторе и генераторе. Даже это кажется привлекательным, но многие сомневаются в его пользе. Часто мы поддерживаем соотношение один к одному. Но некоторые исследователи также проверяют соотношение 5 итераций дискриминатора на одно обновление генератора. Также предлагается сбалансировать обе сети с помощью динамической механики. Но только в последние годы мы получили некоторую поддержку.
С другой стороны, некоторые исследователи оспаривают осуществимость и желательность балансировки этих сетей. Хорошо обученный дискриминатор в любом случае дает генератору качественную обратную связь. Кроме того, непросто научить генератор всегда догонять дискриминатор. Вместо этого мы можем обратить внимание на поиск функции стоимости, которая не имеет градиента, близкого к нулю, когда генератор работает плохо.
Тем не менее, проблемы остаются. Внесено много предложений по функциям затрат, и споры о том, что лучше, продолжаются.
Модель дискриминатора обычно сложнее генератора (больше фильтров и слоев), и хороший дискриминатор дает качественную информацию. Во многих приложениях GAN мы можем столкнуться с узкими местами, где увеличение мощности генератора не приводит к улучшению качества. Пока мы не выявим узкие места и не устраним их, увеличение мощности генератора не кажется приоритетом для многих разделителей.
BigGAN был опубликован в 2018 году с целью объединить некоторые методы GAN для создания лучших изображений на тот момент. В этом разделе мы изучим некоторые еще не рассмотренные практики.
Больший размер партии
Источник (Чем меньше показатель FID, тем лучше) Увеличение размера партии может привести к значительному снижению FID, как показано выше. При большем размере пакета охвачено больше режимов, что обеспечивает лучшие градиенты для обучения обеих сетей. Но тем не менее, BigGAN сообщает, что модель достигает лучшей производительности за меньшее количество итераций, но впоследствии становится нестабильной и даже рушится.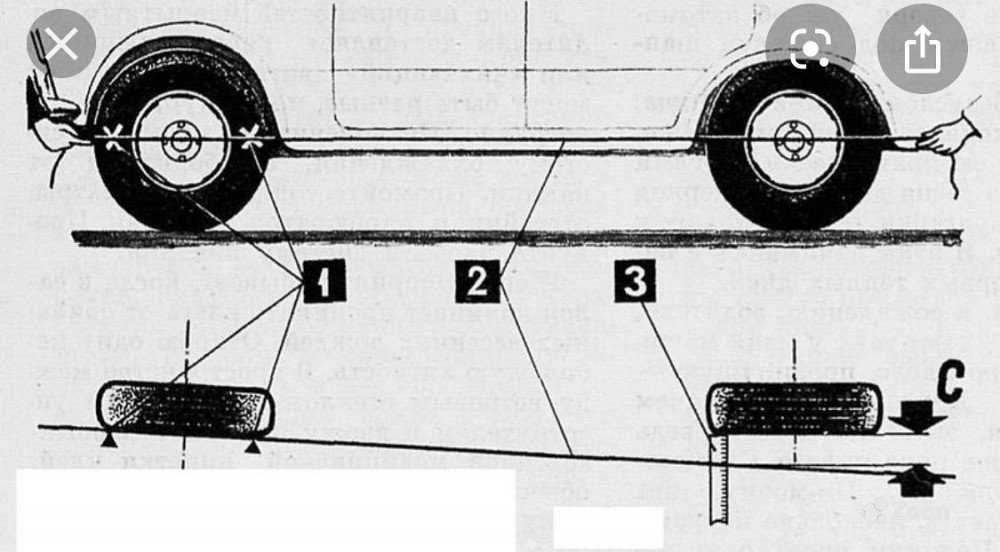 Итак, сохраняйте модель постоянно.
Итак, сохраняйте модель постоянно.
Трюк с усечением
Область низкой плотности вероятности в скрытом пространстве z может не хватить обучающих данных для точного обучения. Таким образом, при создании изображений мы можем избежать этих областей, чтобы улучшить качество изображения за счет изменения. то есть качество изображений повысится, но сгенерированные изображения будут иметь меньшую вариативность стиля. Существуют различные способы усечения входного скрытого пространства z . Общий принцип заключается в том, что когда значения выходят за пределы диапазона, они будут пересчитаны или сжаты до области с более высокой вероятностью.
Увеличение емкости модели
Во время настройки рассмотрите возможность увеличения емкости модели, особенно для слоев с высоким пространственным разрешением. Многие модели показывают улучшение, когда удваивают традиционную емкость, использовавшуюся в то время. Но не делайте этого слишком рано, не проверив дизайн модели и ее реализацию.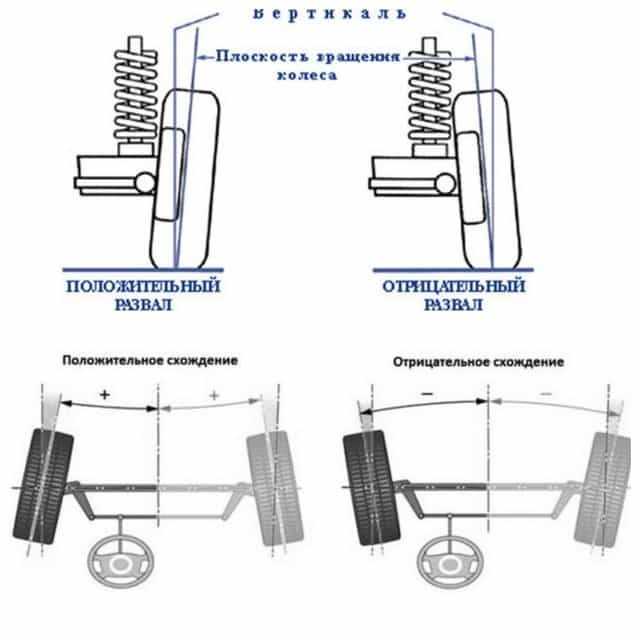
Скользящие средние весов генератора
Веса, используемые генератором, вычисляются из экспоненциального скользящего среднего весов генератора.
Ортогональная регуляризация
Состояние весовой матрицы является хорошо изученной темой. Это исследование того, насколько чувствителен вывод функции к изменениям ее ввода. Это оказывает большое влияние на стабильность тренировок. Матрица Q является ортогональной, если
Если мы умножим x на ортогональную матрицу, изменения в x не будут увеличены. Такое поведение очень желательно для поддержания численной стабильности.
С другими свойствами сохранение ортогональных свойств весовой матрицы может быть привлекательным в глубоком обучении. Мы можем добавить ортогональную регуляризацию, чтобы поощрять такие свойства во время обучения. Это наказывает систему, если Q отклоняется от ортогональной матрицы.
Тем не менее, известно, что это слишком ограничивает, и поэтому BigGAN использует модифицированный термин:
Ортогональная регуляризация также позволяет трюку с усечением быть более успешным в разных моделях.
Инициализация ортогонального веса
Вес модели инициализируется как случайная ортогональная матрица.
Соединение Skip-z
В ванильном GAN скрытый фактор z вводится только для первого слоя. С подключением skip-z, прямым подключением skip (skip-z) из латентного фактора z подключен к нескольким слоям генератора, а не только к первому слою.
В этой статье мы не будем подробно рассматривать улучшение с помощью функции затрат. Это важная тема, и мы рекомендуем читателям прочитать следующую статью:
GAN — всесторонний обзор гангстеров GAN (Часть 2)
В этой статье рассматриваются мотивация и направление исследований GAN по улучшению GAN. Просмотрев их на…
medium.com
Чтобы узнать больше о крутых приложениях GAN:
GAN — Несколько крутых приложений GAN.
Мы добились впечатляющих успехов в первые несколько лет разработки GAN. Больше никаких изображений лица размером с марку, как эти…
medium.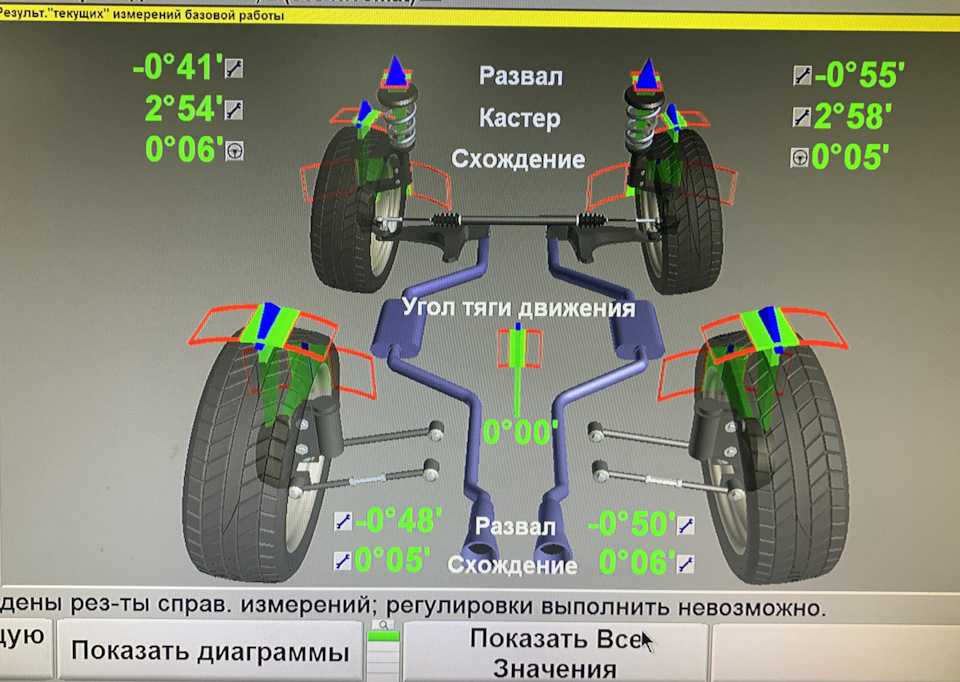 com
com
Все статьи этой серии.
GAN — Серия GAN (от начала до конца)
Полный список наших статей охватывает применение GAN, проблемы и решения.
medium.com
Улучшенные методы обучения GAN
Исследование, финансируемое НАСА: индустриальная цивилизация движется к «необратимому коллапсу»? | Окружающая среда
Новое исследование, частично спонсируемое Центром космических полетов имени Годдарда НАСА, выявило перспективу того, что глобальная индустриальная цивилизация может рухнуть в ближайшие десятилетия из-за неустойчивой эксплуатации ресурсов и все более неравного распределения богатства.
Отмечая, что предупреждения о «крахе» часто кажутся второстепенными или противоречивыми, в исследовании делается попытка осмыслить убедительные исторические данные, показывающие, что «процесс подъема и падения на самом деле является повторяющимся циклом, встречающимся на протяжении всей истории». Случаи серьезного разрушения цивилизации из-за «стремительного краха, часто длящегося столетиями, были довольно обычным явлением».
Независимый исследовательский проект основан на новой междисциплинарной модели «Динамическая динамика человека и природы» (HANDY), которую возглавляет математик-прикладник Сафа Мотешаррей из Национального научного фонда США, поддерживаемого Национальным центром социально-экологического синтеза, в сотрудничестве с команда ученых-естественников и социологов. Модель HANDY создавалась на небольшой грант NASA, но исследование на ее основе проводилось самостоятельно. Исследование, основанное на модели HANDY, было принято к публикации в рецензируемом журнале Elsevier Ecological Economics.
Согласно историческим данным, даже развитые, сложные цивилизации подвержены краху, что ставит под сомнение устойчивость современной цивилизации:
«Падение Римской империи и столь же (если не более) развитые империи Хань, Маурьев и Гупта, а также множество развитых месопотамских империй — все это свидетельствует о том, что передовые, изощренные, сложные и творческие цивилизации могут быть как хрупкими, так и непостоянными».

Исследуя динамику человеческой природы в этих прошлых случаях краха, проект выявляет наиболее важные взаимосвязанные факторы, которые объясняют упадок цивилизации и могут помочь определить риск краха сегодня: а именно: население, климат, вода, сельское хозяйство, и энергия.
Эти факторы могут привести к коллапсу, когда они сходятся, чтобы создать две важные социальные характеристики: «растягивание ресурсов из-за нагрузки на экологическую несущую способность»; и «экономическое расслоение общества на элиты [богатых] и массы (или «простолюдинов») [бедных]». Эти социальные явления играли «центральную роль в характере или в процессе краха» во всех подобных случаях за «последние пять тысяч лет».
В настоящее время высокий уровень экономического расслоения напрямую связан с чрезмерным потреблением ресурсов, причем «элиты», базирующиеся в основном в промышленно развитых странах, несут ответственность за оба эти явления:
«… накопленный излишек не распределяется равномерно по всему обществу, а скорее контролируется элитой.
Масса населения, производя богатство, получает от элиты лишь малую часть его, обычно на уровне или чуть выше прожиточного минимума».

Исследование бросает вызов тем, кто утверждает, что технологии решат эти проблемы за счет повышения эффективности:
«Технологические изменения могут повысить эффективность использования ресурсов, но они также имеют тенденцию к увеличению как потребления ресурсов на душу населения, так и масштабов добычи ресурсов, так что в отсутствие политических эффектов увеличение потребления часто компенсирует повышение эффективности использования ресурсов. .»
Повышение производительности в сельском хозяйстве и промышленности за последние два столетия произошло за счет «увеличения (а не уменьшения) пропускной способности ресурсов», несмотря на резкое повышение эффективности за тот же период.
Моделируя ряд различных сценариев, Мотешаррей и его коллеги пришли к выводу, что в условиях, «точно отражающих реальность сегодняшнего мира.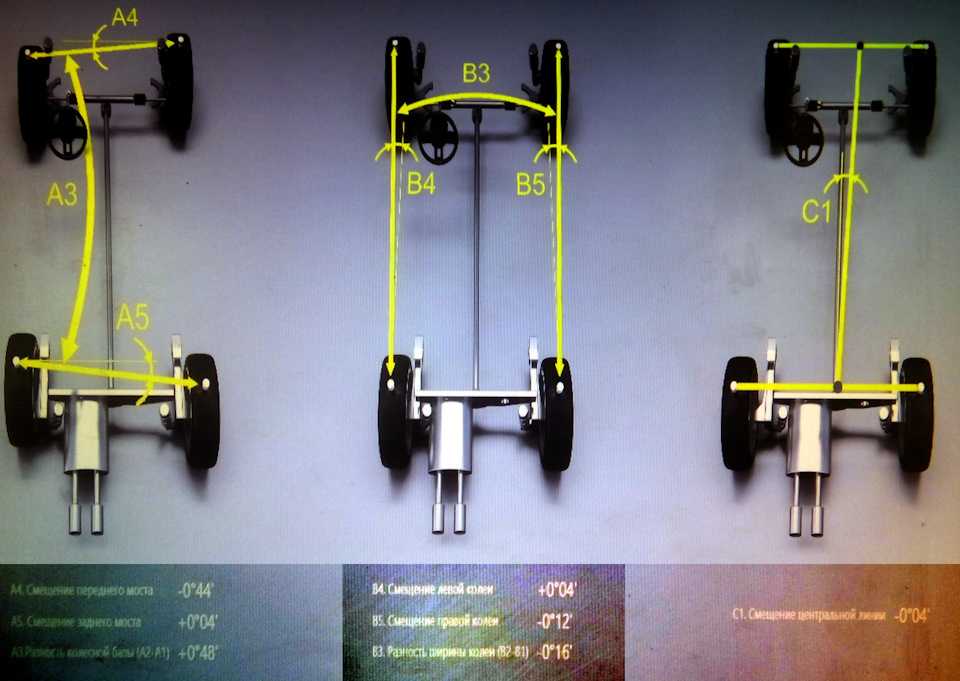
«… кажется, что он находится на устойчивом пути в течение довольно долгого времени, но даже используя оптимальную скорость истощения и начиная с очень небольшого числа элит, элиты в конечном итоге потребляют слишком много, что приводит к голоду среди простолюдинов, который в конечном итоге вызывает крах общества. Важно отметить, что этот крах типа L происходит из-за голода, вызванного неравенством, который вызывает потерю рабочих, а не из-за краха природы ».
Другой сценарий фокусируется на роли продолжающейся эксплуатации ресурсов, обнаруживая, что «при большей скорости истощения упадок простолюдинов происходит быстрее, в то время как элиты все еще процветают, но в конечном итоге простолюдины полностью исчезают, а за ними следуют элиты».
В обоих сценариях элитные монополии на богатство означают, что они защищены от наиболее «пагубных последствий экологического коллапса гораздо позже, чем простолюдины», что позволяет им «продолжать «обычный бизнес», несмотря на надвигающуюся катастрофу». Они утверждают, что тот же самый механизм может объяснить, как «элиты допустили исторические коллапсы, которые, по-видимому, не замечают катастрофической траектории (наиболее ярко проявляющейся в случаях римлян и майя)».
Они утверждают, что тот же самый механизм может объяснить, как «элиты допустили исторические коллапсы, которые, по-видимому, не замечают катастрофической траектории (наиболее ярко проявляющейся в случаях римлян и майя)».
Применяя этот урок к нашему современному затруднительному положению, исследование предупреждает, что:
«В то время как некоторые члены общества могут бить тревогу о том, что система движется к надвигающемуся коллапсу, и поэтому выступают за структурные изменения в обществе, чтобы избежать этого, элиты и их сторонники, выступавшие против внесения этих изменений, могли указать на долгосрочную устойчивую траектория «пока» в поддержку ничегонеделания».
Однако ученые отмечают, что наихудшие сценарии ни в коем случае не являются неизбежными, и предполагают, что соответствующие политические и структурные изменения могут предотвратить крах, если не проложить путь к более стабильной цивилизации.
Два ключевых решения — уменьшить экономическое неравенство, чтобы обеспечить более справедливое распределение ресурсов, и резко сократить потребление ресурсов, полагаясь на менее интенсивные возобновляемые ресурсы и уменьшая прирост населения:
«Коллапса можно избежать, и население может достичь равновесия, если скорость истощения природы на душу населения будет снижена до приемлемого уровня и если ресурсы будут распределяться разумно справедливо».