
LADA Granta с 2011, ремонт антиблокировочной системы инструкция онлайн
Главная / Каталог / Lada / LADA Granta / ВАЗ 2190 с 2011 года, книга по ремонту и каталог деталей в электронном виде / …
Показать содержание книги
Обычно пользователи нашего сайта находят эту страницу по следующим запросам:
нет тормозов ВАЗ Гранта, прокачка тормозов ВАЗ Гранта, схема тормозной системы ВАЗ Гранта, ремонт тормозной системы ВАЗ Гранта, неисправности тормозной системы ВАЗ Гранта, нет тормозов ВАЗ 2190, прокачка тормозов ВАЗ 2190, схема тормозной системы ВАЗ 2190, ремонт тормозной системы ВАЗ 2190, неисправности тормозной системы ВАЗ 2190, нет тормозов LADA 2190, прокачка тормозов LADA 2190, схема тормозной системы LADA 2190, ремонт тормозной системы LADA 2190, неисправности тормозной системы LADA 2190, нет тормозов LADA Гранта, прокачка тормозов LADA Гранта, схема тормозной системы LADA Гранта, ремонт тормозной системы LADA Гранта, неисправности тормозной системы LADA Гранта
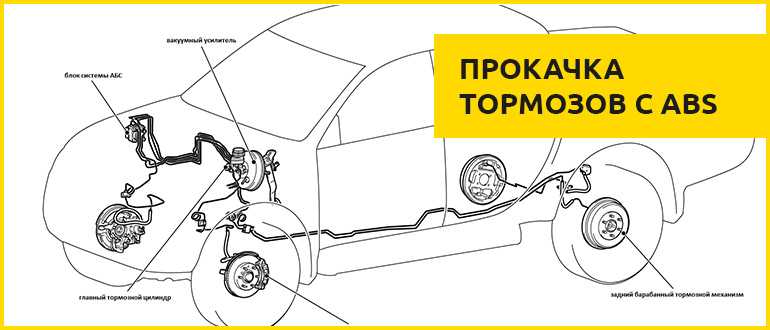
Антиблокировочная система тормозов (ABS) – снятие и установка основных узлов
Внимание
При демонтаже гидроагрегата и трубопроводов принять меры по исключению разлива тормозной жидкости.
Общее описание системы, особенности устройства и работы
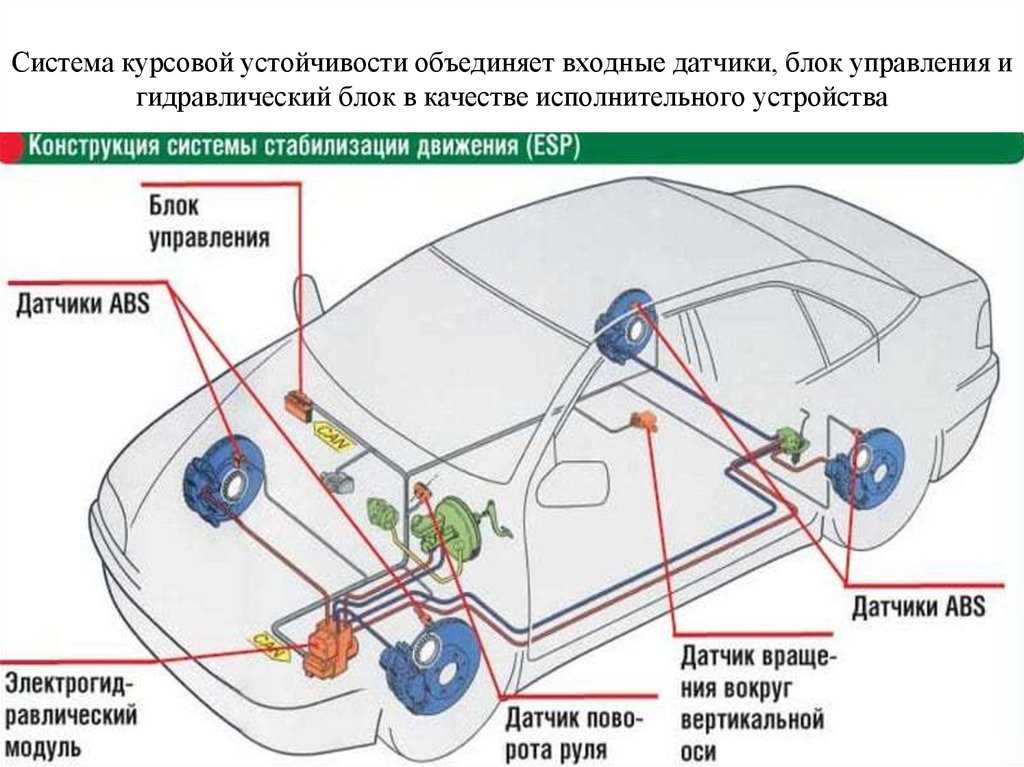
Антиблокировочная система тормозов (ABS) является частью рабочей тормозной системы автомобиля и предназначена для автоматического регулирования степени проскальзывания колес в направлении их вращения во время торможения за счет изменения давления тормозной жидкости в рабочих тормозных цилиндрах с целью предотвращения потери управляемости и устойчивости автомобиля и повышения эффективности торможения.
ABS состоит из следующих основных узлов: — гидроагрегата; — двух датчиков скорости передних колёc — двух датчиков скорости задних колёс; — двух роторов передних колёс, ротор переднего колеса входит в состав наружного шарнира; — двух роторов задних колёс. Гидроагрегат (ГА) конструктивно состоит из электронного блока управления (ЭБУ) и гидромодулятора, содержащего электромагнитные клапаны (ЭМК), возвратный насос и электродвигатель возвратного насоса (ЭВН). Датчики скорости колёс (ДСК) вырабатывают сигналы о скорости каждого колеса автомобиля, которые передаются в электронный блок управления гидроагрегата.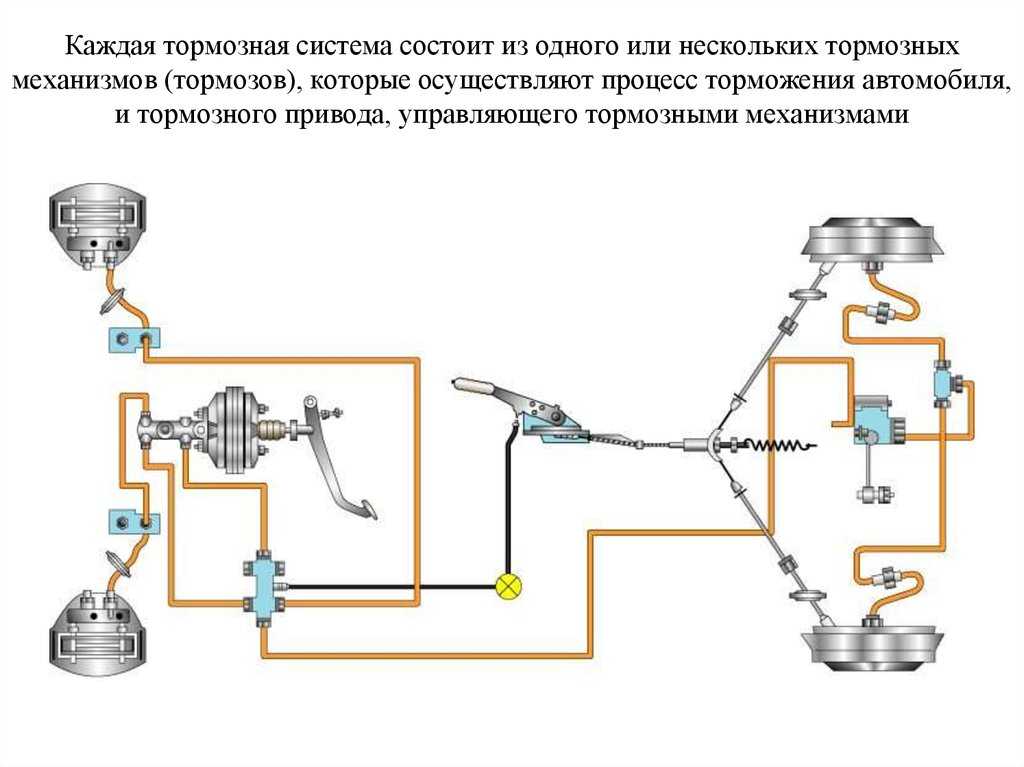 В основу работы колесных датчиков положен принцип электромагнитной индукции. При вращении колеса мимо датчика проходят зубцы и впадины специального ротора и наводят в обмотке датчика электрический сигнал, частота которого пропорциональна уг-ловой скорости колеса и количеству зубцов на роторе. Электронный блок управления производит логическую обработку сигналов о ско-рости колес и, в зависимости от их состояния (чрезмерное ускорение или замедление ко-леса), направляет управляющие команды к гидромодулятору. Гидромодулятор по полученным командам, включая или отключая электромагнит-ные клапаны, снижает, повышает или удерживает постоянным давление тормозной жид-кости в колесных тормозных цилиндрах, обеспечивая тем самым оптимальное регулиро-вание тормозных сил. При снижении давления излишняя тормозная жидкость перекачи-вается возвратным насосом в главный тормозной цилиндр.
В основу работы колесных датчиков положен принцип электромагнитной индукции. При вращении колеса мимо датчика проходят зубцы и впадины специального ротора и наводят в обмотке датчика электрический сигнал, частота которого пропорциональна уг-ловой скорости колеса и количеству зубцов на роторе. Электронный блок управления производит логическую обработку сигналов о ско-рости колес и, в зависимости от их состояния (чрезмерное ускорение или замедление ко-леса), направляет управляющие команды к гидромодулятору. Гидромодулятор по полученным командам, включая или отключая электромагнит-ные клапаны, снижает, повышает или удерживает постоянным давление тормозной жид-кости в колесных тормозных цилиндрах, обеспечивая тем самым оптимальное регулиро-вание тормозных сил. При снижении давления излишняя тормозная жидкость перекачи-вается возвратным насосом в главный тормозной цилиндр.
Гидроагрегат антиблокировочной системы тормозов – снятие и установка
Снятие
Установить автомобиль на двухстоечный подъемник, затормозить стояночным тормозом и выключить зажигание (подъемник электрогидравлический типа П-3,2Г грузо-подъемностью 3,2 т).
Тормозная система Лада Гранта: устройство и работа
Тормоза Лада Гранта необходимы для контроля скорости машины. Они отвечают за плавное снижение скорости до того момента, как автомобиль остановится. Основная задача тормозной системы состоит в том, чтобы обеспечить безопасность людей, которые находятся в салоне автомобиля.
Лада Гранта появилась на отечественных дорогах приблизительно в конце 2011 года. Автомобиль относится к бюджетному классу. Надо сказать, что это достаточно надежная и неприхотливая в обслуживании машина. У нее хорошая управляемость и экономный расход топлива.
Разработана Гранта на базе Калины. Изначально автомобиль выпускался в кузове седан. Начиная с 2013 года был создан хетчбэк. Существует три основные комплектации Гранты: стандартная, нормальная и люксовая. В комплектации «Стандарт» автомобиль оснащается силовым агрегатом в 1,6 л объемом, мощность его составляет 80-90 лошадиных сил. МКПП — пятиступенчатая. Автомобиль имеет передний привод.
Комплектация тормозной системы Гранты
По сравнению с Ладой Калиной Гранта имеет более «продвинутую» тормозную систему. Некоторые автомобили оснащаются системой АБС. В таком случае используется схема тормозов, которые имеют четыре канала. При этом соединение каналов осуществляется по диагональной схеме.
Основным исполнительным элементом антиблокировочной системы является гидромодулятор.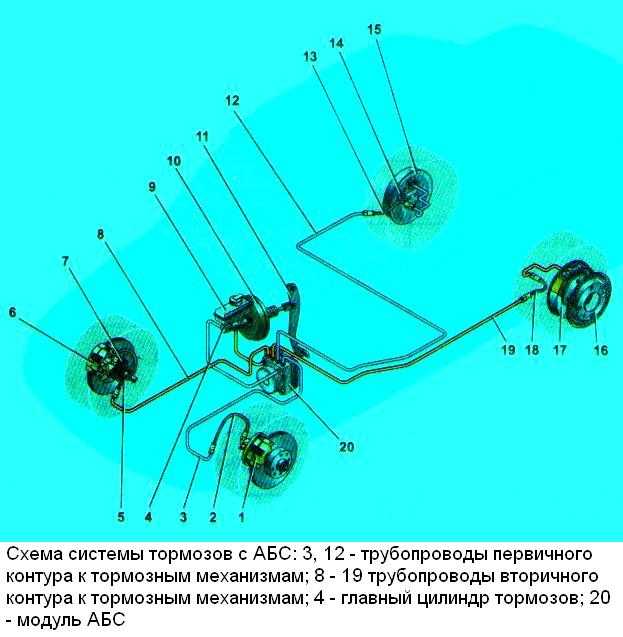 Он представляет собой сложную конструкцию, в которую входит гидронасос, а также клапаны электромагнитного типа. Его монтаж осуществляется в отсеке, в котором располагается мотор. Управляется гидромодулятор при помощи электроники. Передние и задние тормоза оснащаются датчиками скорости вращения колес. Работой тормозов Лада Гранта управляют две системы: рабочая и стояночная. Первая имеет следующее устройство:
Он представляет собой сложную конструкцию, в которую входит гидронасос, а также клапаны электромагнитного типа. Его монтаж осуществляется в отсеке, в котором располагается мотор. Управляется гидромодулятор при помощи электроники. Передние и задние тормоза оснащаются датчиками скорости вращения колес. Работой тормозов Лада Гранта управляют две системы: рабочая и стояночная. Первая имеет следующее устройство:
- Состоит из двух контуров диагонального типа.
Для фиксации автомобиля в одном положении предусмотрено два тормозных контура
- Оснащается гидравлическим приводом.
- Главный тормозной цилиндр оснащен усилителем вакуумного типа.
- Имеется четыре тормозных узла.
- Регулятор давления жидкости находится в задних дисках.
- Передняя тормозная система оснащена дисковым вентилируемым механизмом.
- Задние тормоза относятся к барабанному типу.
Управление колесами при торможении происходит по диагонали — одно переднее и одно заднее.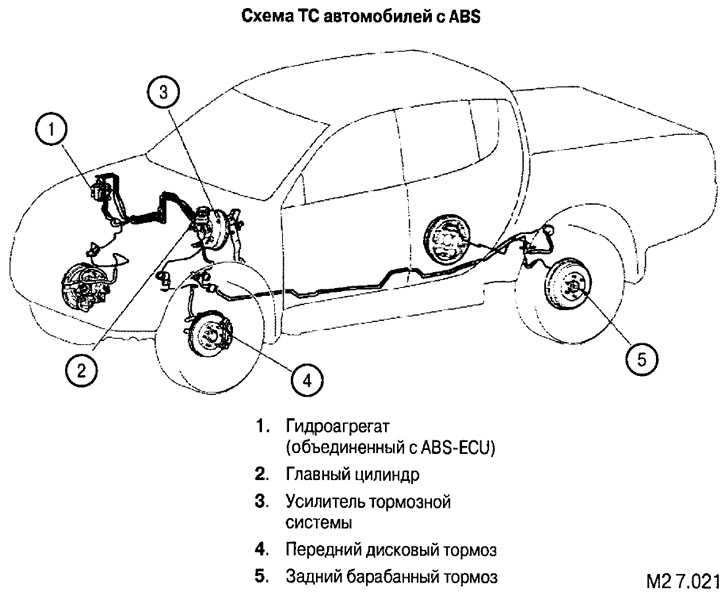 Конструкция контура состоит из механизмов торможения, которые располагаются на переднем правом и заднем левом колесе. Если произошла поломка в одном из контуров, второй контур сможет остановить машину.
Конструкция контура состоит из механизмов торможения, которые располагаются на переднем правом и заднем левом колесе. Если произошла поломка в одном из контуров, второй контур сможет остановить машину.
Для ограничения поступления жидкости в рабочие механизмы, расположенные на задней подвеске, используется регулятор давления жидкости. Когда задняя ось имеет недостаточную нагрузку, при резком торможении это позволяет избежать заноса задней части Лады Гранты. Конструкция исключает возможность блокировки заднего моста.
Корпус регулятора оснащен отверстием, которое закрывается заглушкой из пластмассы. Если происходит подтекание масла, это является признаком того, что нарушена герметизация колец, которые расположены в регуляторе. Вакуумный усилитель предназначен для того, чтобы уменьшить силу надавливания на педаль, которая приводит в действие устройство торможения. Принцип его работы состоит в разрежении, которое возникает в трубопроводе впускного типа во время работы силового агрегата.
Корпус главного цилиндра рабочей системы оснащен бачком, в который заливается жидкость. В свою очередь, крышка бачка оснащается датчиком, который контролирует недостаточный уровень жидкости. Когда уровень жидкости в бачке доходит до критического, на панели приборов загорается контрольная лампочка.
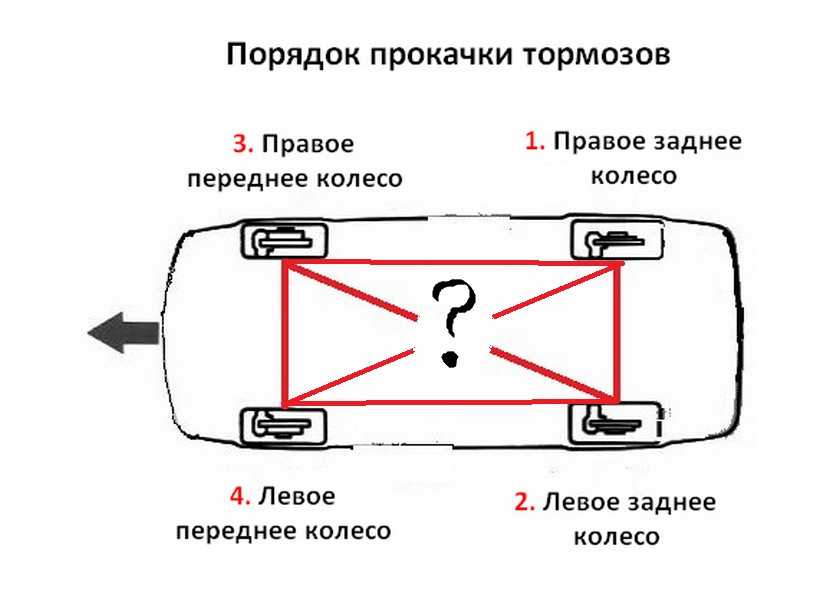
Проверка и прокачка тормозов
Для того чтобы система работала нормально, состояние тормозных колодок рекомендуется проверять не реже, чем раз в месяц. Тормоза в автомобиле Лада Гранта могут приводиться в действие при помощи ручного механизма. Он запускает блокировку задних колес при помощи развода колодок в их барабанах.
Для того чтобы безопасность управления автомобилем была на нужном уровне, следует выполнять проверку и прокачку тормозов.
Проверка выполняется после того, как автомобиль подняли на домкрате. После этого производится демонтаж колес. Затем снимают барабан. Если штифты находятся в открученном состоянии, необходимо произвести ослабление троса ручного тормоза. После осмотра устройства торможения производится смазка и сборка в обратном порядке.
После этого производится демонтаж колес. Затем снимают барабан. Если штифты находятся в открученном состоянии, необходимо произвести ослабление троса ручного тормоза. После осмотра устройства торможения производится смазка и сборка в обратном порядке.
После ремонта в любом из узлов устройства торможения автомобиля Лада Гранта следует произвести прокачку. Выполнять эту работу нужно вдвоем. При прокачке тормозов один человек находится в салоне автомобиля, а второй нажимает до пяти раз педаль тормоза. В это время помощник должен ослабить винт, который находится рядом с диском. Это позволяет спустить жидкость в устройство торможения. Проделывается эта процедура несколько раз. Таким же образом производят регулировку передних тормозов.
Внеклеточные цистеины C226 и C232 опосредуют зависимое от сероводорода ингибирование Orai3-опосредованного поступления кальция в депо
1. Prakriya M, Lewis RS. Депо-управляемые кальциевые каналы. Физиол Преподобный
95: 1383–1436, 2015. doi: 10.1152/physrev.00020.2014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Prakriya M, Lewis RS. Депо-управляемые кальциевые каналы. Физиол Преподобный
95: 1383–1436, 2015. doi: 10.1152/physrev.00020.2014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3. Luik RM, Wu MM, Buchanan J, Lewis RS. Элементарная единица входа Ca 2+ , управляемого депо: локальная активация каналов CRAC с помощью STIM1 в местах соединения ER-плазматической мембраны. Джей Селл Биол 174: 815–825, 2006. doi: 10.1083/jcb.200604015. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
4. Prakriya M, Feske S, Gwack Y, Srikanth S, Rao A, Hogan PG. Orai1 является важной поровой субъединицей канала CRAC. Природа 443: 230–233, 2006. doi: 10.1038/nature05122. [PubMed] [CrossRef] [Академия Google]
5. Соболофф Дж., Спасова М.А., Тан XD, Хевавитарана Т., Сюй В., Гилл Д.Л. Orai1 и STIM восстанавливают функцию депо-управляемых кальциевых каналов. J Биол Хим
281: 20661–20665, 2006. doi: 10.1074/jbc.C600126200. [PubMed] [CrossRef] [Google Scholar]
Соболофф Дж., Спасова М.А., Тан XD, Хевавитарана Т., Сюй В., Гилл Д.Л. Orai1 и STIM восстанавливают функцию депо-управляемых кальциевых каналов. J Биол Хим
281: 20661–20665, 2006. doi: 10.1074/jbc.C600126200. [PubMed] [CrossRef] [Google Scholar]
6. DeHaven WI, Smyth JT, Boyles RR, Putney JW Jr.. Ингибирование кальция и усиление действия кальция в кальциевых каналах Orai1, Orai2 и Orai3, активируемых высвобождением кальция. J Биол Хим 282: 17548–17556, 2007. doi: 10.1074/jbc.M611374200. [PubMed] [CrossRef] [Академия Google]
7. Frischauf I, Muik M, Derler I, Bergsmann J, Fahrner M, Schindl R, Groschner K, Romanin C. Молекулярные детерминанты связи между каналами STIM1 и Orai: дифференциальная активация каналов Orai1-3 спиральным STIM1 -спиральный мутант. J Биол Хим 284: 21696–21706, 2009. doi: 10.1074/jbc.M109.018408. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Frischauf I, Schindl R, Bergsmann J, Derler I, Fahrner M, Muik M, Fritsch R, Lackner B, Groschner K, Romanin C.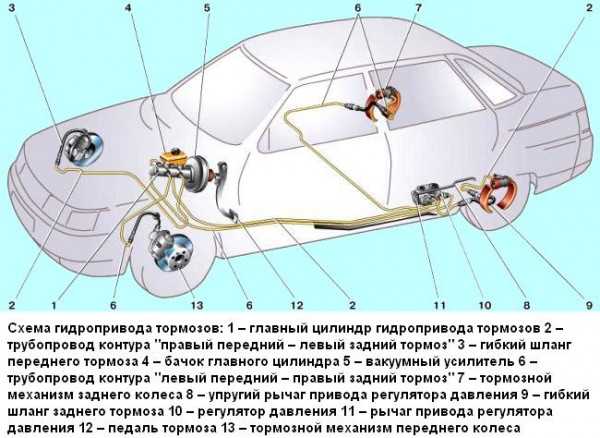 Кооперативность цитозольных доменов Orai настраивает гейтирование, специфичное для подтипа. J Биол Хим
286: 8577–8584, 2011. doi: 10.1074/jbc.M110.187179. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Кооперативность цитозольных доменов Orai настраивает гейтирование, специфичное для подтипа. J Биол Хим
286: 8577–8584, 2011. doi: 10.1074/jbc.M110.187179. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Lee KP, Yuan JP, Zeng W, So I, Worley PF, Muallem S. Молекулярные детерминанты быстрой Ca 2+ -зависимой инактивации и стробирование каналов Орай. Proc Natl Acad Sci USA 106: 14687–14692, 2009. doi: 10.1073/pnas.0904664106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Yoast RE, Emrich SM, Zhang X, Xin P, Johnson MT, Fike AJ, Walter V, Hempel N, Yule DI, Sneyd J, Gill DL, Trebak M. Трио нативных каналов ORAI лежит в основе разнообразия событий передачи сигналов Ca2+. Нац Коммуна 11: 2444, 2020. doi: 10.1038/s41467-020-16232-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Zhang X, Xin P, Yoast RE, Emrich SM, Johnson MT, Pathak T, Benson JC, Azimi I, Gill DL, Monteith GR, Trebak M. Различные фармакологические профили каналов ORAI1, ORAI2 и ORAI3.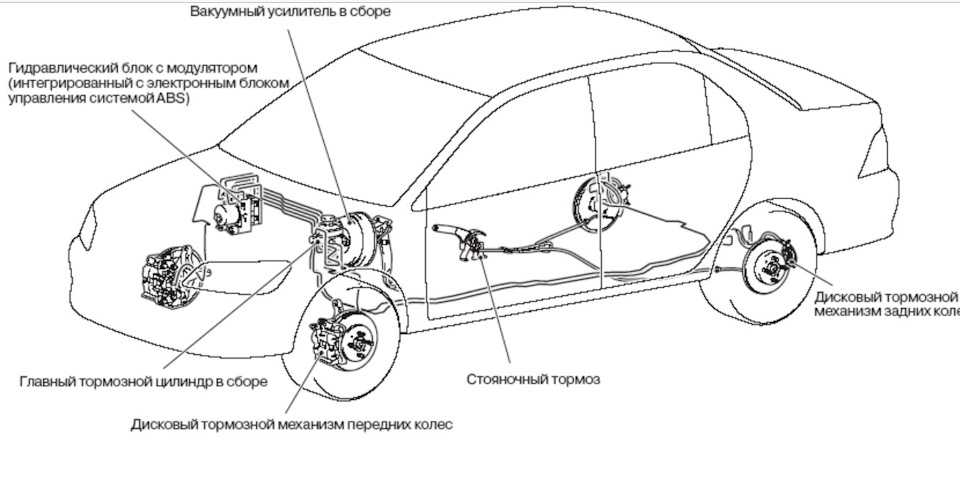 Клеточный кальций
91: 102281, 2020. doi: 10.1016/j.ceca.2020.102281. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Клеточный кальций
91: 102281, 2020. doi: 10.1016/j.ceca.2020.102281. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Bogeski I, Kummerow C, Al-Ansary D, Schwarz EC, Koehler R, Kozai D, Takahashi N, Peinelt C, Griesemer D, Bozem M , Мори Ю., Хот М., Нимейер Б.А. Дифференциальная окислительно-восстановительная регуляция ионных каналов ORAI: механизм настройки передачи сигналов кальция в клетке. Научный сигнал 3: ra24, 2010. doi: 10.1126/scisignal.2000672. [PubMed] [CrossRef] [Академия Google]
13. Kawasaki T, Ueyama T, Lange I, Feske S, Saito N. Индуцированное протеинкиназой C фосфорилирование Orai1 регулирует внутриклеточный уровень Ca 2+ через депо-управляемый канал Ca 2+ . J Биол Хим 285: 25720–25730, 2010. doi: 10.1074/jbc.M109.022996. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Schwindling C, Quintana A, Krause E, Hoth M. Расположение митохондрий контролирует локальный приток кальция в Т-клетки.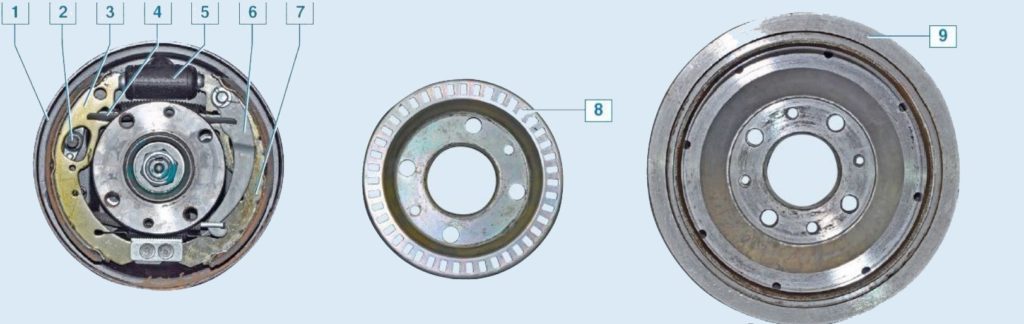 Дж Иммунол
184: 184–190, 2010. doi: 10.4049./jиммунол.0902872. [PubMed] [CrossRef] [Google Scholar]
Дж Иммунол
184: 184–190, 2010. doi: 10.4049./jиммунол.0902872. [PubMed] [CrossRef] [Google Scholar]
15. Скримджер Н.Р., Уилсон Д.П., Рычков Г.Ю. Glu 106 в поре Orai1 способствует быстрой Ca 2+ -зависимой инактивации и pH-зависимости высвобождения Ca 2+ , активированного Ca 2+ (CRAC). Биохим Дж 441: 743–753, 2012. doi: 10.1042/BJ20110558. [PubMed] [CrossRef] [Google Scholar]
16. Sharma A, Ramena G, Yin Y, Premkumar L, Elble RC. CLCA2 является положительным регулятором депо-управляемого входа кальция и TMEM16A. PLoS Один 13: e0196512, 2018. doi: 10.1371/journal.pone.0196512. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
17. Шрикант С., Юнг Х.Дж., Ким К.Д., Суда П., Уайтлеге Дж., Гвак Ю. Новый белок EF-руки, CRACR2A, представляет собой цитозольный кальций. Сенсор 2+ , стабилизирующий каналы CRAC в Т-клетках. Нат клеточный биол 12: 436–446, 2010. doi: 10.1038/ncb2045. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Xiao B, Coste B, Mathur J, Patapoutian A. Зависимая от температуры активация STIM1 индуцирует Ca 2+ приток и модулирует экспрессию генов. Нат Хим Биол
7: 351–358, 2011. doi: 10.1038/nchembio.558. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Xiao B, Coste B, Mathur J, Patapoutian A. Зависимая от температуры активация STIM1 индуцирует Ca 2+ приток и модулирует экспрессию генов. Нат Хим Биол
7: 351–358, 2011. doi: 10.1038/nchembio.558. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
19. Yang Z, Yan H, Dai W, Jing J, Yang Y, Mahajan S, Zhou Y, Li W, Macaubas C, Mellins ED, Shih CC, Fitzpatrick JAJ, Faccio R. Tmem178 отрицательно регулирует депо-управляемый вход кальция в миелоидные клетки посредством ассоциации со STIM1. J Аутоиммунный 101: 94–108, 2019. doi: 10.1016/j.jaut.2019.04.015. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Velmurugan GV, Huang H, Sun H, Candela J, Jaiswal MK, Beaman KD, Yamashita M, Prakriya M, White C. Истощение H 2 S во время ожирения увеличивает поступление в магазин Ca 2+ в макрофагах жировой ткани для увеличения продукции цитокинов. Научный сигнал 8: ra128, 2015. doi: 10.1126/scisignal.aac7135. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Cuevasanta E, Lange M, Bonanata J, Coitiño EL, Ferrer-Sueta G, Filipovic MR, Alvarez B. Реакция сероводорода с дисульфидом и сульфеновой кислотой. кислоты с образованием сильно нуклеофильного персульфида. J Биол Хим
290: 26866–26880, 2015. doi: 10.1074/jbc.M115.672816. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Cuevasanta E, Lange M, Bonanata J, Coitiño EL, Ferrer-Sueta G, Filipovic MR, Alvarez B. Реакция сероводорода с дисульфидом и сульфеновой кислотой. кислоты с образованием сильно нуклеофильного персульфида. J Биол Хим
290: 26866–26880, 2015. doi: 10.1074/jbc.M115.672816. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Francoleon NE, Carrington SJ, Fukuto JM. Реакция H 2 S с окисленными тиолами: образование персульфидов и последствия для биологии H 2 S. Арх Биохим Биофиз 516: 146–153, 2011. doi: 10.1016/j.abb.2011.09.015. [PubMed] [CrossRef] [Google Scholar]
23. Ядав П.К., Мартинов М., Витвицкий В., Серавалли Дж., Ведманн Р., Филипович М.Р., Банерджи Р. Биосинтез и реактивность персульфидов цистеина в передаче сигналов. J Am Chem Soc 138: 289–299, 2016. doi: 10.1021/jacs.5b10494. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Абэ К., Кимура Х. Возможная роль сероводорода как эндогенного нейромодулятора. Джей Нейроски
16: 1066–1071, 1996. doi: 10.1523/JNEUROSCI.16-03-01066.1996. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Джей Нейроски
16: 1066–1071, 1996. doi: 10.1523/JNEUROSCI.16-03-01066.1996. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
25. Коллуру Г.К., Бир С.К., Юань С., Шен С., Пардью С., Ван Р., Кевиль С.Г. Цистатионин-γ-лиаза регулирует артериогенез посредством NO-зависимого рекрутирования моноцитов. Кардиовасц Рес 107: 590–600, 2015. doi: 10.1093/cvr/cvv198. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Li L, Salto-Tellez M, Tan C-H, Whiteman M, Moore PK. GYY4137, новая молекула, высвобождающая сероводород, защищает крыс от эндотоксического шока. Свободный Радик Биол Мед 47: 103–113, 2009. doi: 10.1016/j.freeradbiomed.2009.04.014. [PubMed] [CrossRef] [Google Scholar]
27. Уайтман М., Ли Л., Роуз П., Тан С-Х, Паркинсон Д.Б., Мур П.К. Влияние доноров сероводорода на индуцированное липополисахаридами образование медиаторов воспаления в макрофагах. Антиоксидный окислительно-восстановительный сигнал
12: 1147–1154, 2010. doi: 10.1089./арс. 2009.2899. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2009.2899. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Алансары Д., Богески И., Нимейер Б.А. Упрощение таргетинга Orai3 и функции магазина, управляемой Orai1. Биохим Биофиз Акта 1853: 1541–1550, 2015. doi: 10.1016/j.bbamcr.2015.03.007. [PubMed] [CrossRef] [Google Scholar]
29. Bergsmann J, Derler I, Muik M, Frischauf I, Fahrner M, Pollheimer P, Schwarzinger C, Gruber HJ, Groschner K, Romanin C. Молекулярные детерминанты в пределах N-конца Белок Orai3, контролирующий активацию и управление каналами. J Биол Хим 286: 31565–31575, 2011. doi: 10.1074/jbc.M111.227546. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Томпсон Дж.Л., Шаттлворт Т.Дж. Молекулярная основа активации регулируемого арахидонатом канала Ca 2+ (ARC), депо-независимого канала Orai, с помощью плазматической мембраны STIM1. Дж Физиол 591: 3507–3523, 2013. doi: 10.1113/jphysiol.2013.256784. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Zal T, Gascoigne NRJ. Визуализация эффективности FRET живых клеток с коррекцией фотообесцвечивания. Биофиз Дж
86: 3923–3939, 2004.
[Ошибка в Биофиз J 87: 2915, 2004]. doi: 10.1529/biophysj.103.022087. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Zal T, Gascoigne NRJ. Визуализация эффективности FRET живых клеток с коррекцией фотообесцвечивания. Биофиз Дж
86: 3923–3939, 2004.
[Ошибка в Биофиз J 87: 2915, 2004]. doi: 10.1529/biophysj.103.022087. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Zhou Y, Wang Y, Gill DL. Оценка молекулярной природы интерфейса связи STIM1/Orai1 с использованием подходов FRET. В: Каналы входа кальция в невозбудимых клетках, под редакцией Козак Дж. А., Патни Дж. В. мл. Бока-Ратон, Флорида: CRC Press/Taylor & Francis, 2018. [PubMed] [Google Scholar]
33. Kaminski CF, Rees EJ, Schierle GSK. Количественный протокол для визуализации FRET живых клеток на основе интенсивности. Методы Мол Биол 1076: 445–454, 2014. doi: 10.1007/9.78-1-62703-649-8_19. [PubMed] [CrossRef] [Google Scholar]
34. Bird GS, DeHaven WI, Smyth JT, Putney JW Jr.. Методы изучения депо-управляемого поступления кальция. Методы 46: 204–212, 2008. doi: 10.1016/j.ymeth.2008.09.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. DeHaven WI, Smyth JT, Boyles RR, Bird GS, Putney JW Jr..
Комплексное действие 2-аминоэтилдифенилбората на депо кальция. J Биол Хим
283: 19265–19273, 2008. doi: 10.1074/jbc.M801535200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
DeHaven WI, Smyth JT, Boyles RR, Bird GS, Putney JW Jr..
Комплексное действие 2-аминоэтилдифенилбората на депо кальция. J Биол Хим
283: 19265–19273, 2008. doi: 10.1074/jbc.M801535200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Чжан С.Л., Козак Дж.А., Цзян В., Еромин А.В., Чен Дж., Ю.Ю., Пенна А., Шен В., Чи В., Кахалан М.Д. Депо-зависимый и -независимый режимы, регулирующие высвобождение Ca 2+ , активируют активность канала Ca 2+ Orai1 и Orai3 человека. J Биол Хим 283: 17662–17671, 2008. doi: 10.1074/jbc.M801536200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Барр В.А., Бернот К.М., Шрикант С., Гвак И., Балагопалан Л., Риган К.К., Хелман Д.Дж., Соммерс К.Л., О-Хора М., Рао А. , Самельсон Л.Е. Динамическое движение кальциевого сенсора STIM1 и кальциевого канала Orai1 в активированных Т-клетках: точки и дистальные колпачки. Мол Биол Селл 19: 2802–2817, 2008. doi: 10.1091/mbc.e08-02-0146. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Lis A, Peinelt C, Beck A, Parvez S, Monteilh-Zoller M, Fleig A, Penner R. CRACM1, CRACM2 и CRACM3 сохранены. -управляемые каналы Ca 2+ с различными функциональными свойствами. Карр Биол
17: 794–800, 2007. doi: 10.1016/j.cub.2007.03.065. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Lis A, Peinelt C, Beck A, Parvez S, Monteilh-Zoller M, Fleig A, Penner R. CRACM1, CRACM2 и CRACM3 сохранены. -управляемые каналы Ca 2+ с различными функциональными свойствами. Карр Биол
17: 794–800, 2007. doi: 10.1016/j.cub.2007.03.065. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Vaeth M, Yang J, Yamashita M, Zee I, Eckstein M, Knosp C, Kaufmann U, Karoly Jani P, Lacruz RS, Flockerzi V, Качковиц И., Пракрия М., Феске С. ORAI2 модулирует вход кальция в депо и опосредованный Т-клетками иммунитет. Нац Коммуна 8: 14714, 2017. doi: 10.1038/ncomms14714. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
40. Миньен О., Томпсон Дж.Л., Шаттлворт Т.Дж. Молекулярная архитектура регулируемого арахидонатом Ca 2+ -селективного канала ARC представляет собой пентамерную сборку субъединиц Orai1 и Orai3. Дж Физиол 587: 4181–4197, 2009. doi: 10.1113/jphysiol.2009.174193. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41. Zhang X, Zhang W, González-Cobos JC, Jardin I, Romanin C, Matrougui K, Trebak M. Комплексная роль STIM1 в активации независимые от магазина каналы Orai1/3. J Ген Физиол
143: 345–359, 2014. doi: 10.1085/jgp.201311084. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Zhang X, Zhang W, González-Cobos JC, Jardin I, Romanin C, Matrougui K, Trebak M. Комплексная роль STIM1 в активации независимые от магазина каналы Orai1/3. J Ген Физиол
143: 345–359, 2014. doi: 10.1085/jgp.201311084. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Powell CR, Dillon KM, Matson JB. Обзор доноров сероводорода (H 2 S): химия и потенциальное терапевтическое применение. Биохим Фармакол 149: 110–123, 2018. doi: 10.1016/j.bcp.2017.11.014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
43. Benchoam D, Cuevasanta E, Möller MN, Alvarez B. Окисление сероводорода и персульфидов биологически значимыми окислителями. Антиоксиданты (Базель) 8: 48, 2019. doi: 10.3390/antiox8020048. [PMC бесплатная статья] [PubMed] [CrossRef] [Google Scholar]
44. Jiang B, Tang G, Cao K, Wu L, Wang R. Молекулярный механизм H 2 S-индуцированная активация K ATP каналы. Антиоксидный окислительно-восстановительный сигнал
12: 1167–1178, 2010. doi: 10.1089/ars.2009.2894. [PubMed] [CrossRef] [Google Scholar]
doi: 10.1089/ars.2009.2894. [PubMed] [CrossRef] [Google Scholar]
45. Ma SF, Luo Y, Ding YJ, Chen Y, Pu SX, Wu HJ, Wang ZF, Tao BB, Wang WW, Zhu YC. Сероводород нацеливается на мотив Cys320/Cys529 в Kv4.2, чтобы ингибировать калиевые каналы Ito в кардиомиоцитах и регулирует фатальную аритмию при инфаркте миокарда. Антиоксидный окислительно-восстановительный сигнал 23: 129–147, 2015. doi: 10.1089/ars.2014.6094. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Ge S-N, Zhao MM, Wu DD, Chen Y, Wang Y, Zhu J-H, Cai W-J, Zhu Y-Z, Zhu Y-C. Сероводород нацеливается на остатки EGFR Cys797/Cys798, вызывая эндоцитоз Na + /K + -АТФазы и ингибирование эпителиальных клеток почечных канальцев, а также увеличивает экскрецию натрия у крыс с хронической солевой нагрузкой. Антиоксидный окислительно-восстановительный сигнал 21: 2061–2082, 2014. doi: 10.1089/ars.2013.5304. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Тао Б.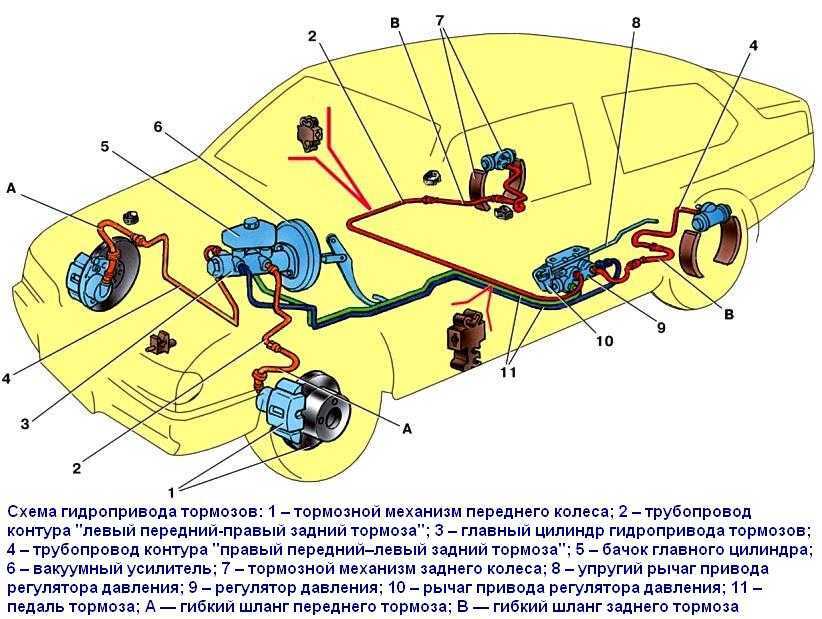 Б., Лю С.Ю., Чжан С.К., Фу В., Цай В.Дж., Ван Ю., Шэнь Ц., Ван М.Дж., Чен Ю., Чжан Л.Дж., Чжу Ю.З., Чжу Ю.К. VEGFR2 функционирует как H 2 S-нацеливающая рецепторная протеинкиназа с его новой дисульфидной связью Cys1045-Cys1024, служащей в качестве специфического молекулярного переключателя действия сероводорода в эндотелиальных клетках сосудов. Антиоксидный окислительно-восстановительный сигнал
19: 448–464, 2013. doi: 10.1089/ars.2012.4565. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Б., Лю С.Ю., Чжан С.К., Фу В., Цай В.Дж., Ван Ю., Шэнь Ц., Ван М.Дж., Чен Ю., Чжан Л.Дж., Чжу Ю.З., Чжу Ю.К. VEGFR2 функционирует как H 2 S-нацеливающая рецепторная протеинкиназа с его новой дисульфидной связью Cys1045-Cys1024, служащей в качестве специфического молекулярного переключателя действия сероводорода в эндотелиальных клетках сосудов. Антиоксидный окислительно-восстановительный сигнал
19: 448–464, 2013. doi: 10.1089/ars.2012.4565. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Наварро-Борелли Л., Сомасундарам А., Ямашита М., Рен Д., Миллер Р.Дж., Пракрия М. Взаимодействия STIM1-Orai1 и конформационные изменения Orai1, выявленные вживую -клеточная FRET-микроскопия. Дж Физиол 586: 5383–5401, 2008. doi: 10.1113/jphysiol.2008.162503. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Парк С.И., Гувер П.Дж., Маллинз Ф.М., Баххават П., Ковингтон Э.Д., Раунсер С., Вальц Т., Гарсия К.С., Долметш Р.Э., Льюис Р.С. STIM1 кластеризуется и активирует каналы CRAC посредством прямого связывания цитозольного домена с Orai1.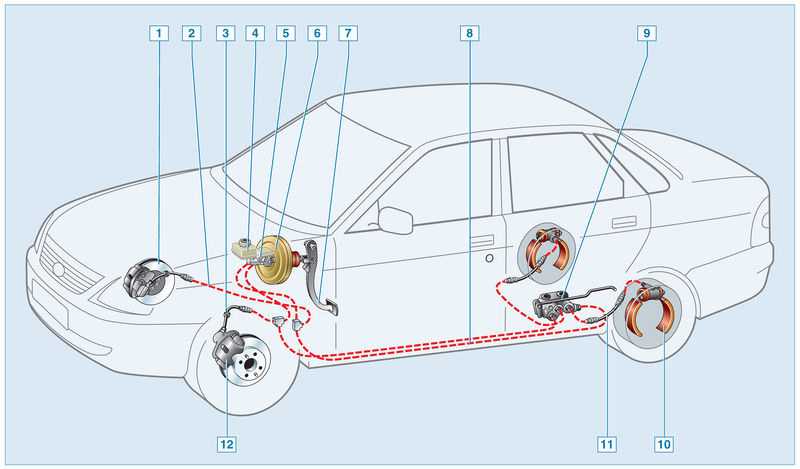 Клетка
136: 876–890, 2009. doi: 10.1016/j.cell.2009.02.014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Клетка
136: 876–890, 2009. doi: 10.1016/j.cell.2009.02.014. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Алансари Д., Шмидт Б., Дёрр К., Богески И., Ригер Х., Клесс А., Нимейер Б.А. Тиолзависимая внутримолекулярная блокировка каналов Orai1. научный представитель 6: 33347, 2016. doi: 10.1038/srep33347. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Yazbeck P, Tauseef M, Kruse K, Amin MR, Sheikh R, Feske S, Komarova Y, Mehta D. Фосфорилирование STIM1 по Y361 рекрутирует Orai1 в точку STIM1 и индуцирует проникновение Ca 2+ . научный представитель 7: 42758, 2017. doi: 10.1038/srep42758. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Emrich SM, Yoast RE, Xin P, Arige V, Wagner LE, Hempel N, Gill DL, Sneyd J, Yule DI, Trebak M. Omnitemporal хореография всех пяти STIM/Orai и IP3R лежит в основе сложности Ca 9 млекопитающих.0007 2+ сигнализация. Представитель ячейки
34: 108760, 2021. doi: 10.1016/j. celrep.2021.108760. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
celrep.2021.108760. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Cao X, Ding L, Xie Z-Z, Yang Y, Whiteman M, Moore PK, Bian J-S. Обзор синтеза, метаболизма и измерения сероводорода: является ли модулирование сероводорода новым терапевтическим средством против рака? Антиоксидный окислительно-восстановительный сигнал 31: 1–38, 2019. doi: 10.1089/ars.2017.7058. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
54. Li Z, Polhemus DJ, Lefer DJ. Эволюция сероводородной терапии для лечения сердечно-сосудистых заболеваний. Цирк Рез 123: 590–600, 2018. doi: 10.1161/CIRCRESAHA.118.311134. [PubMed] [CrossRef] [Google Scholar]
55. Jia J, Li J, Cheng J. H 2 Лечение ишемического инсульта на основе S: возможности и проблемы. Инсульт Васк Нейрол 4: 63–66, 2019. doi: 10.1136/svn-2018-000194. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
56. Zhang Y, Tang Z-H, Ren Z, Qu SL, Liu M-H, Liu LS, Jiang Z-S. Сероводород, следующее мощное профилактическое и лечебное средство при старении и возрастных заболеваниях.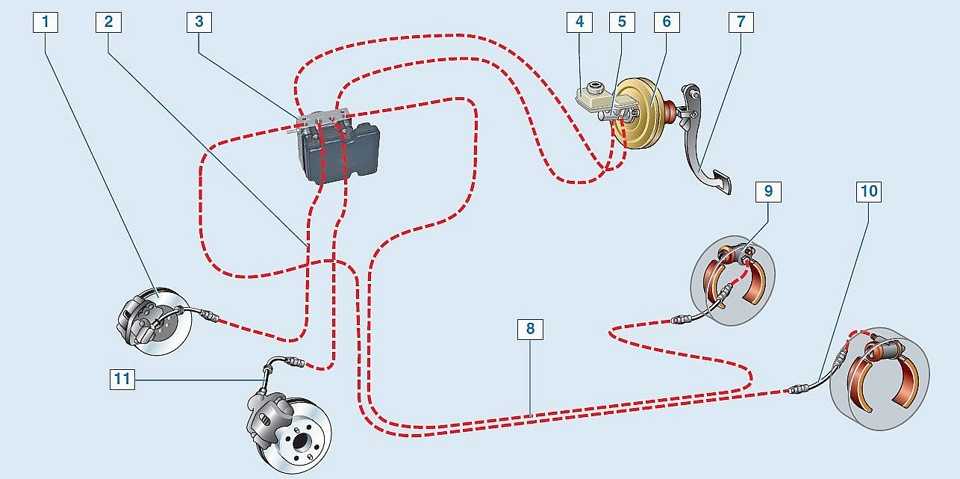 Мол Селл Биол
33: 1104–1113, 2013. doi: 10.1128/MCB.01215-12. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Мол Селл Биол
33: 1104–1113, 2013. doi: 10.1128/MCB.01215-12. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
57. Азими И., Милевский М.Дж.Г., Чалмерс С.Б., Япа КТДС, Робитайл М., Генри С., Бейлли Г.Дж., Томпсон Э.В., Робертс-Томсон С.Дж., Монтейт Г.Р. ORAI1 и ORAI3 в молекулярных подтипах рака молочной железы и идентификация ORAI3 как чувствительного к гипоксии гена и регулятора реакции на гипоксию. Раки (Базель) 11: 208, 2019. doi: 10.3390/cancers11020208. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
58. Benzerdjeb N, Sevestre H, Ahidouch A, Ouadid-Ahidouch H. Orai3 является прогностическим маркером метастазирования и выживаемости при резектабельной аденокарциноме легкого. Онкотаргет 7: 81588–81597, 2016. doi: 10.18632/oncotarget.13149. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
59. Hasna J, Hague F, Rodat-Despoix L, Geerts D, Leroy C, Tulasne D, Ouadid-Ahidouch H, Kischel P. Orai3 кальциевый канал и устойчивость к химиотерапии в клетках рака молочной железы: соединение p53.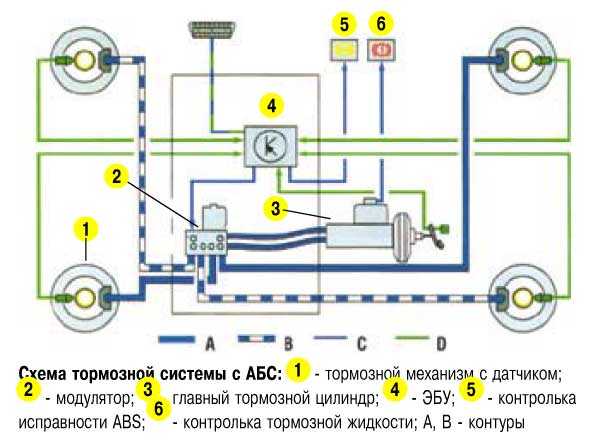 Смерть клеток
25: 693–707, 2018. doi: 10.1038/s41418-017-0007-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Смерть клеток
25: 693–707, 2018. doi: 10.1038/s41418-017-0007-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
60. Holzmann C, Kilch T, Kappel S, Armbrüster A, Jung V, Stöckle M, Bogeski I, Schwarz EC, Peinelt C. ICRAC контролирует скорость андрогенный ответ в первичных эпителиальных клетках предстательной железы человека и изменяется при раке предстательной железы. Онкотаргет 4: 2096–2107, 2013. doi: 10.18632/oncotarget.1483. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
61. Мотиани Р.К., Абдуллаев И.Ф., Требак М. Новый нативный депо-управляемый кальциевый канал, кодируемый Orai3: селективная потребность Orai3 по сравнению с Orai1 в рецепторе эстрогена. положительные по сравнению с отрицательными по рецептору эстрогена клетками рака молочной железы. J Биол Хим 285: 19173–19183, 2010. doi: 10.1074/jbc.M110.102582. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
62. Saliba Y, Keck M, Marchand A, Atassi F, Ouillé A, Cazorla O, Trebak M, Pavoine C, Lacampagne A, Hulot J-S, Farès Н.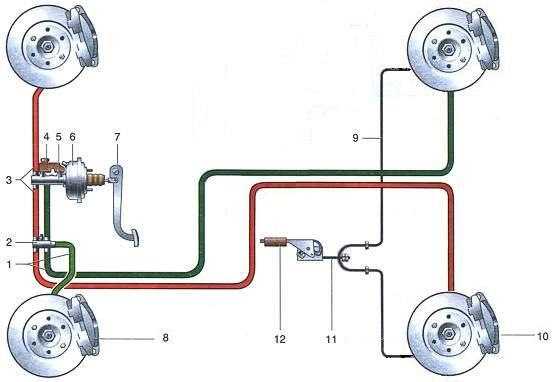 , Фоконье Дж., Ломпре А.М. Возникновение активности Orai3 при сердечной гипертрофии. Кардиовасц Рес
105: 248–259, 2015. doi: 10.1093/cvr/cvu207. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Фоконье Дж., Ломпре А.М. Возникновение активности Orai3 при сердечной гипертрофии. Кардиовасц Рес
105: 248–259, 2015. doi: 10.1093/cvr/cvu207. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. González-Cobos JC, Zhang X, Zhang W, Ruhle B, Motiani RK, Schindl R, Muik M, Spinelli AM, Bisaillon JM, Shinde AV , Fahrner M, Singer HA, Matrougui K, Barroso M, Romanin C, Trebak M. Store-независимые каналы Orai1/3, активируемые интракринным лейкотриеном C4: роль в неоинтимальной гиперплазии. Цирк Рез 112: 1013–1025, 2013. doi: 10.1161/CIRCRESAHA.111.300220. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Im Onlineshop kaufen ▷ bequem & sicher |
- Zertifiziertes Umweltmanagement
- Производство и дизайн сделано в Германии
- Markenqualität seit 1958
- Сихерес Айнкауфен
Добро пожаловать в мир d-c-fix ®
МАГАЗИН
- США – Амазонка
БЛОГ И ВДОХНОВЕНИЕ
Wenn im Handumdrehen aus der abgenutzten Kommode wieder ein echtes Decor-Highlight wird, die in die Jahre gekommene Duschkabine im neuen Look erstrahlt und die rustikale Küche modern pledrkwigtligel anderen Fliesens haben unsere Produkte von d-c-fix® wieder einmal bewiesen, dass große Veränderungen weder teuer noch langwierig sein müssen.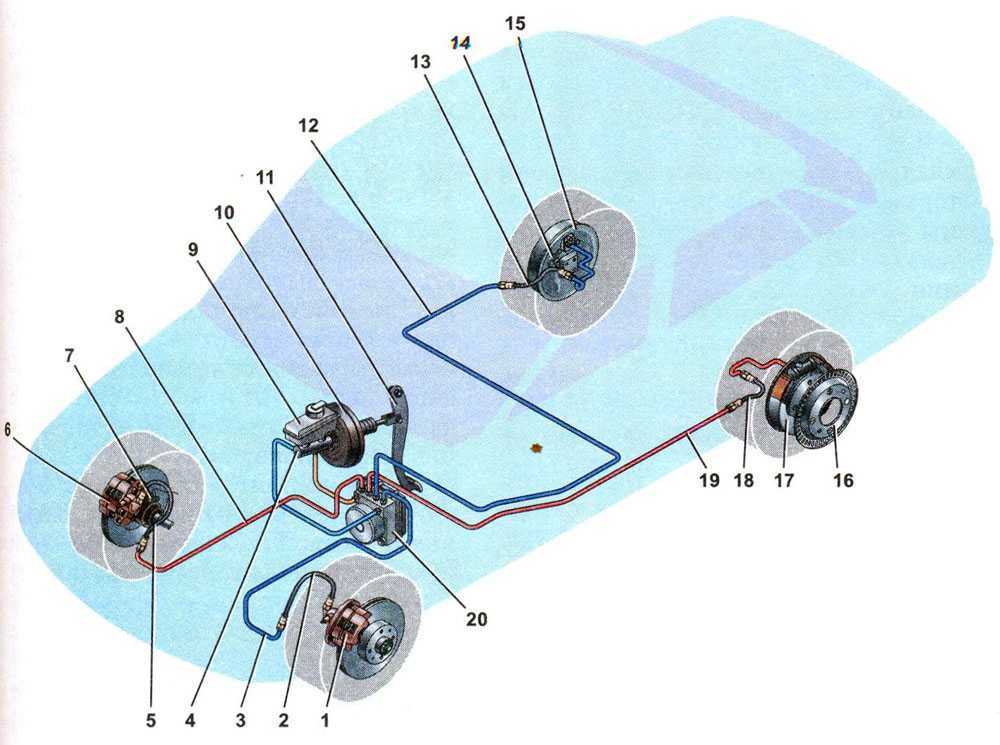 Überzeuge Dich von unseren Dekofolien aller Art сразу в unserem d-c-fix® Onlineshop und erhalte noch mehr Anregungen für künftige Deko-Projekte.
Überzeuge Dich von unseren Dekofolien aller Art сразу в unserem d-c-fix® Onlineshop und erhalte noch mehr Anregungen für künftige Deko-Projekte.
Im Home-Decor-Bereich und bei DIY-Fans mit einer Vorliebe für Interior Design sind unsere vielfältigen Dekofolien bereits Weltweit ebenso bekannt wie geschätzt und eröffnen den Anwendern täglich zahlreiche neue kreative Horizonte. Unser d-c-fix® Sortiment ist sehr umfangreich und umfasst dabei Klebefolien mit einer Vielzahl unterschiedlicher Prints wie Holz- und Steinoptiken, grafischen Mustern sowie ausdrucksstarken Unis – иммерьте ориентацию и актуальные тенденции Trends der Deko-Welt.
Neben den klassischen d-c-fix® Klebefolien zur Verschönerung alter Schrank-Fronten, Tischplatten oder Türen, sind zudem selbstklebende sowie statisch-haftende Fensterfolien Teil unseres Produkt-Programms. Abgerundet wird das Sortiment für Home-Decor-Liebhaber zudem durch selbstklebende Wandbeläge, Bodenfliesen sowie abwischbare Tischdecken & -sets – hier führen wir mittlerweile auch eine große Anzahl PVC-freier Produkte. Klick Dich einfach durch und entdecke sie im offiziellen d-c-fix® Online-Shop.
Klick Dich einfach durch und entdecke sie im offiziellen d-c-fix® Online-Shop.
d-c-fix® Home Decoration online kaufen
Mit unseren selbstklebenden d-c-fix® Folien sind Deiner Kreativität keine Grenzen gesetzt. Mit ihrer Hilfe wirst Du im Handumdrehen zum DIY-Künstler und Home-Decor-Experten und kannst Deine ganz persönlichen Vorstellungen verwirklichen. So wird Dein Zuhause zum Unikat mit Wiedererkennungswert. Du hast Dich schon Lange an der Optik Deines Regals sattgesehen? Dann los und lass Deinem Sinn für Interior Design freien Lauf. Wie wäre es zum Beispiel mit einer Dekofolie в классическом современном Marmor-Optik? Wir versprechen: Dieses Decor kommt nie aus der Mode und passt zu vielen unterschiedlichen Einrichtungsstilen!
Falls Du mit Deiner Einrichtung bereits absolut zufrieden bist, sind vielleicht unsere Glas- und Funktionsfolien etwas für Dich. Die Folien für die Fenster sind z. B. nicht nur eine tolle Möglichkeit, Deine Fensterflächen optisch aufzuwerten – unsere Designs reichen von ornamentalen und grafischen Mustern bis zu Folien mit Milchglas-Effekt – sondern auch einen Eleganten Sichtschutz zu schaffen.