
| Двигатель | ||||
|---|---|---|---|---|
| Рабочий объем (см³) | 2694 | 2755 | 3956 | 2694 |
| Тип двигателя | Бензиновый | Дизельный | Бензиновый | Бензиновый |
| Максимальная мощность | 163 | 200 | 249 | 163 |
| Количество клапанов на цилиндр | 4 | 4 | 4 | 4 |
| Вид топлива | Бензин с октановым числом 91 и выше | Дизельное топливо с цетановым числом не менее 48 | Бензин с октановым числом 95 и выше | Бензин с октановым числом 91 и выше |
| Код двигателя | 2TR-FE | 1GD-FTV | 1GR-FE | 2TR-FE |
| Число и тип расположения цилиндров | 4, Рядное | 6, V-образное | 4, Рядное | |
| Клапанный механизм | DOHC цепной привод с электронной системой изменения фаз газораспределения VVT-I | DOHC цепной привод | DOHC цепной привод с системой изменения фаз газораспределения Dual VVT-I | DOHC цепной привод с электронной системой изменения фаз газораспределения VVT-I |
| Диаметр цилиндра х ход поршня (мм х мм) | 95 x 95 | 92 x 103. 6 6 | 94 x 95 | 95 x 95 |
| Система впрыска топлива | Распределенный впрыск | Система непосредственного впрыска под давлением COMMON RAIL и интеркуллером | Распределенный впрыск | Распределенный впрыск |
| Степень сжатия | 10.2:1 | 15.6:1 | 10.4:1 | 10.2:1 |
| Максимальная мощность (кВт при об/мин) | 120/5200 | 147/3400 | 183/5600 | 120/5200 |
| Максимальный крутящий момент (Нм при об/мин) | 246/3900-3900 | 500/1600-2800 | 381/4400-4400 | 246/3900-3900 |
| Вес | ||||
| Снаряженная масса (кг) | 2105-2265 | 2235-2305 | 2150-2415 | 2105-2265 |
| Максимальная масса (кг) | 2850 | 2990 | 2900 | 2850 |
| Масса буксируемого прицепа, оборудованного тормозами (кг) | 1500 | 3000 | 3000 | 1500 |
| Масса буксируемого прицепа, не оборудованного тормозами (кг) | 750 | 750 | 750 | |
| Максимальная масса автомобиля — на переднюю ось (кг) | 1450 | 1450 | 1450 | 1450 |
| Максимальная масса автомобиля — на заднюю ось (кг) | 1800 | 1800 | 1800 | 1800 |
| Размеры | ||||
| Длина (мм) | 4840 | 4840 | 4840 | 4840 |
| Ширина (мм) | 1885 | 1885 | 1885 | 1885 |
| Высота (мм) | 1895 | 1895 | 1895 | 1895 |
| Колесная база (мм) | 2790 | 2790 | 2790 | 2790 |
| Колея задних колес (мм) | 1605 | 1605 | 1605 | 1605 |
| Колея передних колес (мм) | 1605 | 1605 | 1605 | 1605 |
| Передний свес (мм) | 975 | 975 | 975 | 975 |
| Задний свес (мм) | 1075 | 1075 | 1075 | 1075 |
| Внутренние размеры | ||||
| Количество мест | 5 | 5 | 5 | 5 |
| Потребление топлива | ||||
| Городской цикл (л/100 км) | 13.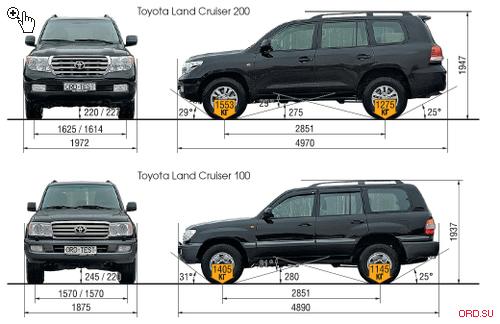 9 9 | 9.5 | 15.5 | 13.9 |
| Экологический класс | Евро 5 | Евро 5 | Евро 5 | Евро 5 |
| Емкость топливного бака (л) | 87 | 87 | 87 | |
| Загородный цикл (л/100 км) | 9.3 | 6.7 | 9.4 | 9.3 |
| Смешанный цикл (л/100 км) | 11 | 7.7 | 11.6 | 11 |
| Трансмиссия | ||||
| Тип привода | Постоянный полный | Постоянный полный | Постоянный полный | Постоянный полный |
| Тип трансмиссии | Механическая | Гидромеханическая | Гидромеханическая | Гидромеханическая |
| Число передач | 5 | 6 | 6 | 6 |
| 1-я передача | 3. 830 830 | 3.600 | 3.600 | 3.600 |
| 2-я передача | 2.062 | 2.090 | 2.090 | 2.090 |
| 3-я передача | 1.436 | 1.488 | 1.488 | 1.488 |
| 4-я передача | 1.000 | 1.000 | 1.000 | 1.000 |
| 5-я передача | 0.838 | 0.687 | 0.687 | 0.687 |
| 6-я передача | 0.580 | 0.580 | 0.580 | 0.580 |
| Передача заднего хода | 4.220 | 3.732 | 3.732 | 3.732 |
| Главная передача | 4.555 | 3.909 | 3.909 | 4.777 |
| Динамические характеристики | ||||
| Максимальная скорость (км/ч) | 165 | 175 | 160 | |
| Время разгона 0-100 км/час (сек) | 13. 8 8 | — | — | 13.9 |
| Колесные диски и шины | ||||
| Размер шин | 245/70 R17 | 265/65 R17 | 265/60 R18 | 265/65 R17 |
| Колесные диски | Стальные | Легкосплавные | Легкосплавные | Легкосплавные |
| Рулевое управление | ||||
| Дополнительные системы | HPS (гидроусилитель руля) | HPS (гидроусилитель руля) | HPS (гидроусилитель руля) | HPS (гидроусилитель руля) |
| Тип рулевого механизма | Рулевой механизм типа «шестерня-рейка» | Рулевой механизм типа «шестерня-рейка» | Рулевой механизм типа «шестерня-рейка» | Рулевой механизм типа «шестерня-рейка» |
| Минимальный радиус разворота – по колесам (м) | 5,8 | 5,8 | 5,8 | 5,8 |
| Передаточное отношение | 15,7 | 15,7 | 15,7 | 15,7 |
| Количество оборотов (между крайними положениями руля) | 3 | 3 | 3 | 3 |
| Тормоза | ||||
| ABS | Антиблокировочная система тормозов | Антиблокировочная система тормозов | Антиблокировочная система тормозов | Антиблокировочная система тормозов |
| Передние тормоза (тип) | Вентилируемые тормозные диски | Вентилируемые тормозные диски | Вентилируемые тормозные диски | Вентилируемые тормозные диски |
| Задние тормоза (тип) | Вентилируемые тормозные диски | Вентилируемые тормозные диски | Вентилируемые тормозные диски | Вентилируемые тормозные диски |
| EBD | Электронная система распределения тормозного усилия | Электронная система распределения тормозного усилия | Электронная система распределения тормозного усилия | Электронная система распределения тормозного усилия |
| BAS | Усилитель экстренного торможения | Усилитель экстренного торможения | Усилитель экстренного торможения | Усилитель экстренного торможения |
| Подвеска | ||||
| Передняя подвеска | Независимая, рычажная, пружинная, с гидравлическими телескопическими а-ми | Независимая, рычажная, пружинная, с гидравлическими телескопическими а-ми | Независимая, рычажная, пружинная, с гидравлическими телескопическими а-ми | Независимая, рычажная, пружинная, с гидравлическими телескопическими а-ми |
| Задняя подвеска | Зависимая, пружинная, с гидравлическими телескопическими амортизаторами | Зависимая, пружинная, с гидравлическими телескопическими амортизаторами | Зависимая, пружинная, с гидравлическими телескопическими амортизаторами | Зависимая, пружинная, с гидравлическими телескопическими амортизаторами |
| Вместимость | ||||
| Длина салона (мм) | 1825 | 1825 | 1825 | 1825 |
| Ширина салона (мм) | 1565 | 1565 | 1565 | 1565 |
| Высота салона (мм) | 1240 | 1240 | 1240 | 1240 |
| Тип кузова | Универсал | Универсал | Универсал | Универсал |
| Объем багажного отделения при поднятых задних сиденьях (л) | 621 | 621 | 621 | 621 |
| Объем багажного отделения при сложенных задних сиденьях (л) | 1934 | 1934 | 1934 | 1934 |
| Эксплуатационные характеристики | ||||
| Дорожный просвет (мм) | 215 | 215 | 215 | 215 |
| Безопасность | ||||
| TRC | Антипробуксовочная система | Активная антипробуксовочная система (A-TRC) | Активная антипробуксовочная система (A-TRC) | Антипробуксовочная система |
| DAC | — | Система помощи при спуске по склону | Система помощи при спуске по склону | — |
| HAC | — | Система помощи при старте на подъеме | Система помощи при старте на подъеме | — |
| VSC | Система курсовой устойчивости | Система курсовой устойчивости | Система курсовой устойчивости | Система курсовой устойчивости |
4. 6 AT 4WD 6 AT 4WD |
4.5 D AT 4WD | |
| Кузов | ||
| Ширина грузового отсека, мм | 1670 | 1670 |
| Колея задних колёс, мм | 1645 | 1645 |
| Колея передних колёс, мм | 1650 | 1650 |
| Количество мест для сидения | 5/7 | 5/7 |
| Минимальный объём багажника, л | 259 | 259 |
| Максимальный объём багажника, л | 1267 | 1267 |
| Диаметр разворота, м | 11.8 | 11.8 |
| Снаряженная масса, кг | 2585 | 2585 |
| Разрешённая масса автопоезда, кг | 6800 | 6800 |
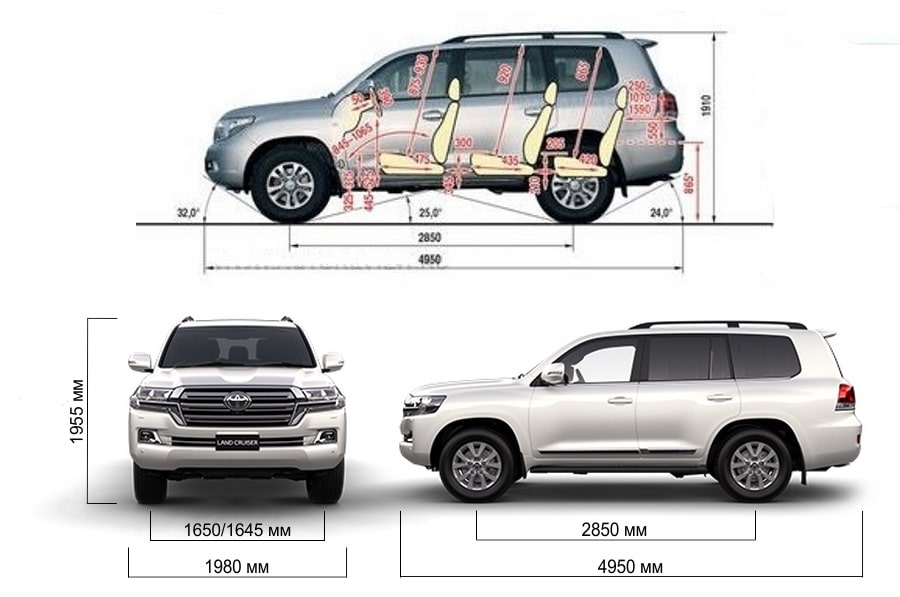
| Колёсная база, мм | 2850 | 2850 |
| Усилитель руля | Гидроусилитель | Гидроусилитель |
| Длина, мм | 4950 | 4950 |
| Ширина, мм | 1980 | 1980 |
| Высота, мм | 1955 | 1955 |
| Дорожный просвет, мм | 225 | 225 |
| Количество дверей | 5 | 5 |
| Объем бензобака, л | 93 | 93 |
| Допустимая полная масса, кг | 3350 | 3350 |
| Двигатель | ||
| Тип двигателя | Бензиновый | Дизельный |
| Объем двигателя в литрах, л | 4. 6 6 |
4.5 |
| Рабочий объем, см3 | 4608 | 4461 |
| Конфигурация | V-образный | V-образный |
| Количество цилиндров | 8 | 8 |
| Количество клапанов на цилиндр | 4 | 4 |
| Диаметр цилиндра, мм | 94 | 86 |
| Длина хода поршня, мм | 83 | 96 |
| Тип впуска | Распределенный впрыск | Непосредственный впрыск |
| Максимальная мощность, л.с. | 309 | 249 |
| Обороты максимальной мощности, мин., об./мин. | 5500 | 2800 |
| Обороты максимальной мощности, макс., об./мин. | Нет данных | 3600 |
| Максимальный крутящий момент | 439 | 650 |
Обороты макс. крут. момента, мин., об./мин. крут. момента, мин., об./мин. |
3400 | 1600 |
| Обороты макс. крут. момента, макс., об./мин. | Нет данных | 2600 |
| Наддув | — | Битурбо |
| Рекомендуемое топливо | АИ-95 | ДТ |
| Интеркулер | — | Да |
| Трансмиссия | ||
| Коробка передач | Автомат | Автомат |
| Количество передач | 6 | 6 |
| Привод | Полный | Полный |
| Ходовая часть | ||
| Передняя подвеска | Независимая, пружинная, рычажная, с гидравлическими телескопическими амортизаторами, со стабилизатором поперечной устойчивости | Независимая, пружинная, рычажная, с гидравлическими телескопическими амортизаторами, со стабилизатором поперечной устойчивости |
| Задняя подвеска | Зависимая пружинная | Независимая, пружинная |
| Диаметр переднего обода, мм | 18 | 18 |
| Ширина профиля передней шины, мм | 285 | 285 |
| Высота профиля передней шины, мм | 60 | 60 |
| Диаметр передней шины, мм | 18 | 18 |
| Диаметр расположения отверстий в диске (PCD), мм | 150 | 150 |
| Количество передних крепёжных отверстий | 5 | 5 |
| Диаметр заднего обода, мм | 18 | 18 |
| Ширина профиля задней шины, мм | 285 | 285 |
| Высота профиля задней шины, мм | 60 | 60 |
| Диаметр задней шины, мм | 18 | 18 |
| Диаметр расположения отверстий в диске (PCD), мм | 150 | 150 |
| Количество задних крепёжных отверстий | 5 | 5 |
| Тормозная система | ||
| Передние тормоза | Дисковые вентилируемые | Дисковые вентилируемые |
| Задние тормоза | Дисковые вентилируемые | Дисковые вентилируемые |
| Динамические характеристики и расход топлива | ||
| Максимальная скорость, км/ч | 205 | 210 |
| Время разгона до 100 км/ч, с | 8. 6 6 |
8.9 |
| Расход топлива в городе, л/100 км | 18.2 | 12 |
| Расход топлива на шоссе, л/100 км | 11.4 | 9.1 |
| Расход топлива в смешанном цикле, л/100 км | 13.9 | 10.2 |
| Объем бензобака, л | 93 | 93 |
Тойота Ленд Крузер 300 (2021-2022)
Издание Best Car раздобыло информацию о новом поколении внедорожника Toyota Land Cruiser 300, премьера которого запланирована на 1 августа 2021 года, а прием заказов в Японии обещают открыть уже весной. Вот какие сведения о грядущей новинке удалось узнать.
Во-первых, новый Тойота Ленд Крузер 300 (2021-2022) останется рамным внедорожником, при этом автомобиль все же получит модернизированную раму и серьезно пересмотренное шасси. Постоянный полный привод разумеется останется на своем месте.
Каталог Toyota
Во-вторых, грядущий Крузак 2021 года окажется несколько крупнее предшественника.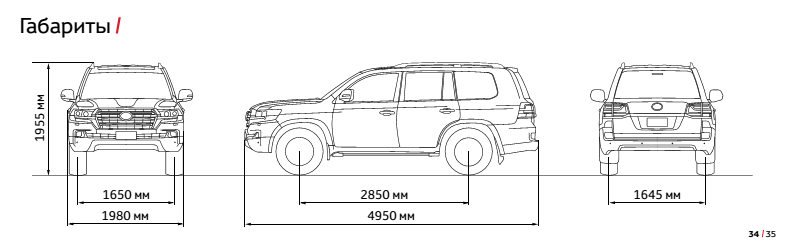 В длину внедорожник будет достигать 4 970 мм (+ 20), в ширину — 1 985 (+ 5), в высоту — 1 870 (- 10). Колесная база автомобиля увеличится до 2 900 мм (+ 50), что позволит повысить стабильность на высоких скоростях.
В длину внедорожник будет достигать 4 970 мм (+ 20), в ширину — 1 985 (+ 5), в высоту — 1 870 (- 10). Колесная база автомобиля увеличится до 2 900 мм (+ 50), что позволит повысить стабильность на высоких скоростях.
В-третьих, внедорожник получит совершенно другие двигатели. Ожидается, что на смену нынешнему 4,6-литровому V8 мощностью 309 л.с. придет 3,5-литровый V6 с двойным наддувом, который ставится на Lexus LS 500. На седане он развивает 421 л.с. (600 Нм) и трудится в паре с 10-диапазонным автоматом, при этом для внедорожника его отдачу могут поднять до 450 сил.
► Подпишитесь на наш канал в Telegram
Также в линейке новой модели Toyota Land Cruiser 300 появится гибридная модификация с 3,5-литровым ДВС на 299 л.с. (356 Нм) и 180-сильным электродвигателем. Альтернативой станет 3,3-литровый дизель, подробности о котором пока не сообщаются.
Долгое время по сети гуляли различные рендеры грядущего внедорожника, а в апреле двадцать первого в сеть утекли фото нового Ленд Крузер 300 без камуфляжа. Как и ожидалось, автомобиль получил эволюционные изменения в дизайне, сохранив пропорции и массивную решетку радиатора. Спереди появились более узкие фары и другой бампер, задние фонари стали компактнее, а угол наклона боковых окошек в районе задних стоек заметно увеличили.
Как и ожидалось, автомобиль получил эволюционные изменения в дизайне, сохранив пропорции и массивную решетку радиатора. Спереди появились более узкие фары и другой бампер, задние фонари стали компактнее, а угол наклона боковых окошек в районе задних стоек заметно увеличили.
Можно не сомневаться, что японцы сделают салон Ленд Крузер 300 более современным и технологичным, в оснащение войдут современная мультимедийная система и множество электронных ассистентов, а чуть позже на его базе дебютирует следующий Лексус LX 570.
Если автомобиль представят осенью 2021-го (ранее сроки постоянно менялись), то старт его продаж в России можно ждать либо к концу года, либо все же в начале 2022-го. Комплектации и цены станут известны позже, при этом стоимость модели наверняка заметно вырастет.
Двигатель 5VZ-FE | Характеристики, проблемы, тюнинг
Характеристики двигателя 5VZ-FE
| Производство | Toyota Motor Manufacturing Alabama |
| Марка двигателя | 5VZ |
| Годы выпуска | 1995-2004 |
| Материал блока цилиндров | чугун |
| Система питания | инжектор |
| Тип | V-образный |
| Количество цилиндров | 6 |
| Клапанов на цилиндр | 4 |
| Ход поршня, мм | 82 |
| Диаметр цилиндра, мм | 93. 5 5 |
| Степень сжатия | 9.6 |
| Объем двигателя, куб.см | 3378 |
| Мощность двигателя, л.с./об.мин | 190/4800 |
| Крутящий момент, Нм/об.мин | 298/3600 |
| Топливо | 95 |
| Экологические нормы | — |
| Вес двигателя, кг | 190 |
| Расход топлива, л/100 км (для 4Runner) — город — трасса — смешан. | 20.0 11.0 14.0 |
| Расход масла, гр./1000 км | до 1000 |
| Масло в двигатель | 0W-30 5W-30 5W-40 10W-30 |
| Сколько масла в двигателе, л | 5.3 |
| Замена масла проводится, км | 7000-10000 |
| Рабочая температура двигателя, град. | ~90 |
| Ресурс двигателя, тыс. км — по данным завода — на практике | — 300+ |
| Тюнинг, л.с. — потенциал — без потери ресурса | 400+ н.  д. д. |
| Двигатель устанавливался | Toyota 4Runner Toyota Land Cruiser Prado Toyota Tacoma Toyota Tundra Toyota T-100 Toyota Granvia ГАЗ 3111 Волга |
| КПП, -5МКПП -4АКПП -5АКПП | Toyota R150F Toyota A340E/F Toyota A650E |
Надежность, проблемы и ремонт двигателя Тойота 5VZ-FE
Очередной двигатель для внедорожных моделей Toyota появился в 1995 году и был разработан на базе 3VZ-E. Блок цилиндров V6 с 60° развалом достался от 3VZE, но цилиндры были увеличены в диаметре с 87.5 мм до 93.5 мм, в них были помещены новые поршни под степень сжатия 9.6. Это позволило добиться рабочего объема в 3.4 литра.
Вместо одновальных ГБЦ, на 5VZ были использованы более современные головки, с двумя распредвалами. Они аналогичны тем, что применены на 3VZ-FE, но с некоторыми изменениями. Распредвалы на 5VZ более низовые, чем на 3-х литровом предшественнике. Также был заменен ремень ГРМ на другой. На 5VZ был использован маслоохладитель, а также доработан вентилятор системы охлаждения. Впускной коллектор был сделан более длинным, чем на 3VZ, для улучшения показателей на низких оборотах, система изменения длины ACIS не была использована. Выпускной коллектор оптимизированный, также несколько изменилась система зажигания и был применен фазированный впрыск топлива (как на 1MZ).
На 5VZ был использован маслоохладитель, а также доработан вентилятор системы охлаждения. Впускной коллектор был сделан более длинным, чем на 3VZ, для улучшения показателей на низких оборотах, система изменения длины ACIS не была использована. Выпускной коллектор оптимизированный, также несколько изменилась система зажигания и был применен фазированный впрыск топлива (как на 1MZ).
Вышеописанные изменения позволили оптимизировать двигатель для внедорожников, пикапов и коммерческой техники.
Для 5VZ-FE, компанией TRD, выпускались компрессор киты, которые позволяли увеличить мощность этих моторов до 245 л.с., а крутящий момент до 360 Нм.
Производство 5VZ было свернуто в 2004 году, а подкапотное пространство тойотовских SUV было плотно оккупировано более свежим 1GR-FE.
Проблемы и недостатки двигателей Тойота 5VZ
Основные проблемы 3.4-х литрового мотора такие же, как на 3-х литровом 3VZ, но проблема с прокладкой ГБЦ менее выражена и сам мотор несколько понадежней. Детальней об этом можно прочитать здесь.
Детальней об этом можно прочитать здесь.
Помимо Тойот, этот мотор использовался на автомобилях ГАЗ Волга, также это популярный вариант для свапа в другие российские авто (УАЗ Буханка, Барс, ГАЗ Газель и прочие).
Тюнинг двигателя Toyota 5VZ-FE
5VZ Компрессор
Имея такой двигатель, лезть в него со спортивными валами, пытаться переделать впуск-выпуск и прочее, не слишком разумное занятие, к тому же жутко дорогое. Для этого мотора можно купить компрессор кит от TRD, на базе компрессора Eaton M62. Новый стоит достаточно немало, но б/у можно найти за приемлемые деньги. Форсунки лучше использовать от 2JZ-GE. На давлении 0.5 бар, на сток поршнях, вы получите около 250-260 л.с. и значительную прибавку крутящего момента во всем диапазоне. Заменив шкив на 56 мм (2.2″), вы увеличите наддув до 0.6 бар и получите около 280 л.с. на 98 бензине. Установив спортивный выхлоп без катализаторов, вы добавите еще около 10 л.с. Можно купить шкив 2.1″ (или вообще 2″), тем самым поднять давление до 0. 75 бар и получить более 300 л.с., но нужно снижать степень сжатия либо ставить впрыск водометанола.
75 бар и получить более 300 л.с., но нужно снижать степень сжатия либо ставить впрыск водометанола.
РЕЙТИНГ ДВИГАТЕЛЯ: 4
<<НАЗАД
2016 Toyota Land Cruiser Prado: новый 2.8 турбодизель, 6АКПП
Последний настоящий.Тест-пилот Дмитрий ШИМАНОВИЧ
Сегодня на тесте — один из немногих, сохранивших верность традициям. Автомобиль, который с гордо поднятой головой несет легендарное имя: Land Cruiser.
За последние пару десятков лет ни один класс автомобилей не изменился столь значительно, как внедорожники. От поколения к поколению они становились комфортнее, мощнее и дружелюбнее к водителю, теряя порой при этом саму суть – внедорожные способности. Но сегодня на тесте — один из немногих, сохранивших верность традициям. Автомобиль, который с гордо поднятой головой несет легендарное имя: Land Cruiser.
Смотреть БОЛЬШОЙ ТЕСТ на нашем канале ЮТУБ:
Внешних отличий и вправду не много. Кроме уже упомянутой тонировки – рейлинги на крыше и светодиодные фары ближнего света. Все. Основная работа велась с технической частью автомобиля. Появилась система стабилизации кузова, гасящая раскачку, много других изменений, призванных повысить комфорт. Главное отличие обновленного автомобиля — абсолютно новая силовая установка. Прежняя связка из архаичной трехлитровой турбодизельной четверки и довольно флегматичного пятиступенчатого «автомата» была отправлена на пенсию. Объем турбодизеля уменьшился до 2,8 литра, а количество передач АКПП достигло шести.
Кроме уже упомянутой тонировки – рейлинги на крыше и светодиодные фары ближнего света. Все. Основная работа велась с технической частью автомобиля. Появилась система стабилизации кузова, гасящая раскачку, много других изменений, призванных повысить комфорт. Главное отличие обновленного автомобиля — абсолютно новая силовая установка. Прежняя связка из архаичной трехлитровой турбодизельной четверки и довольно флегматичного пятиступенчатого «автомата» была отправлена на пенсию. Объем турбодизеля уменьшился до 2,8 литра, а количество передач АКПП достигло шести.
Не смотря на уменьшение объема двигателя, мощность увеличилась. Новый мотор выдает 177 л.с., прибавив 4 лошадки, а крутящий момент и вовсе вырос с 410 до 450 н/м, что особенно важно для внедорожника. Но первое и главное впечатление от нового двигателя – комфорт. Больше никаких вибраций на холостых и заунывного воя нагнетателя. Интенсивный разгон не сопровождается прежним тарахтением, звук мотора стал значительно тише и получил приятный, породистый тембр. Казалось бы, такая малость, ведь ни динамика, ни салон не изменились вообще, а автомобиль сразу ощущается совершенно иначе. АКПП оставила положительные впечатления. Диапазоны в ней меняются ощутимо быстрее, а обороты двигателя на крейсерских скоростях ниже, что является плюсом в копилку экономичности. Ну, а особенно радует тот факт, что, обзаведясь хорошими манерами, Prado не растерял своей сути. Он по-прежнему остается одним из последних представителей вымирающего нынче вида. Он – настоящий, бескомпромиссный 100% внедорожник. В его арсенале две принудительные блокировки и непробиваемая подвеска. У него жесткий задний мост и понижающая передача. Он имеет прекрасно настроенную систему помощи на тяжелом бездорожье и отличную геометрическую проходимость. Он может преодолевать броды до 70 см в глубину и двигаться по уклонам до 45%. А после этого, выбравшись на асфальт, он порадует Вас комфортом бизнес-класса и усладит Ваш слух звучанием 14-ти динамиков аудиосистемы JBL. Prado подскажет водителю о наличии автомобиля в мертвой зоне, поможет припарковаться с помощью камер кругового обзора, сам включит фары и «дворники» и даже охладит Ваши напитки.
Казалось бы, такая малость, ведь ни динамика, ни салон не изменились вообще, а автомобиль сразу ощущается совершенно иначе. АКПП оставила положительные впечатления. Диапазоны в ней меняются ощутимо быстрее, а обороты двигателя на крейсерских скоростях ниже, что является плюсом в копилку экономичности. Ну, а особенно радует тот факт, что, обзаведясь хорошими манерами, Prado не растерял своей сути. Он по-прежнему остается одним из последних представителей вымирающего нынче вида. Он – настоящий, бескомпромиссный 100% внедорожник. В его арсенале две принудительные блокировки и непробиваемая подвеска. У него жесткий задний мост и понижающая передача. Он имеет прекрасно настроенную систему помощи на тяжелом бездорожье и отличную геометрическую проходимость. Он может преодолевать броды до 70 см в глубину и двигаться по уклонам до 45%. А после этого, выбравшись на асфальт, он порадует Вас комфортом бизнес-класса и усладит Ваш слух звучанием 14-ти динамиков аудиосистемы JBL. Prado подскажет водителю о наличии автомобиля в мертвой зоне, поможет припарковаться с помощью камер кругового обзора, сам включит фары и «дворники» и даже охладит Ваши напитки.
Новый мотор 2.8 ТУРБОДИЗЕЛЬ выдает 177 л.с., прибавив 4 лошадки, а крутящий момент и вовсе вырос с 410 до 450 н/м, что особенно важно для внедорожника!
Конечно, он далеко не идеален. Его могли сделать более дружелюбным к водителю. Убрать, к примеру, довольно большие крены, усилив стабилизаторы. Или сделать его мягче. Однако ради этого пришлось бы поступиться артикуляцией подвески и ее энергоемкостью, следовательно, ухудшив внедорожные способности. В эпоху кроссоверов, Prado остается одним из последних настоящих. Солдат, сменивший полевую форму и винтовку на строгий костюм и дорогие часы, но внутри оставшийся честным, надежным боевым товарищем. Он – не подведет. Он – Toyota Land Cruiser Prado.
ТТХ:2.8 турбодизель, 6АКПП
Комплектация «Люкс», 5 мест
Оценочные ПЛЮСЫ и МИНУСЫ:
+ тихий, экономичный дизель;
+ великолепные внедорожные способности;
+ богатое оснащение;
— жесткая подвеска;
— недостаток динамики на высоких скоростях;
— ручка на передней стойке ограничивает обзор.
Тест-пилот Дмитрий ШИМАНОВИЧ
Больше тестов вседорожников и легковых в нашем плэйлисте Toyota на канале АВТОПАНОРАМА в ЮТУБ:
Подписывайтесь на наш канал ЮТУБ
и смотрите первыми
самые свежие тесты и обзоры премьер:
https://www.youtube.com/user/KlimovichKatya
Паджеро 4 или Прадо 150 что лучше на сегодняшний день?
Вечные соперники – автомобильные монстры Тойота и Митсубиши нередко ставят в тупик будущих владельцев, предлагая им самостоятельно выбрать наиболее подходящую внедорожную версию. Поэтому мнение, что лучше Паджеро 4 или Прадо 150, по большей части опирается на субъективную оценку владельцев, имеющих опыт эксплуатации либо одного, либо другого внедорожника.
Официальные представители с обеих сторон не спешат предоставить на общий суд батл-испытания, количество отказов систем узлов и агрегатов, возвратов и доработок у своих автомобилей. Они часто преподносят сухую информацию в виде ТТХ. Выдают желаемое за действительное, а иногда просто кратко описывают те или иные функции каждого авто, еще больше запутывая будущих владельцев.
Они часто преподносят сухую информацию в виде ТТХ. Выдают желаемое за действительное, а иногда просто кратко описывают те или иные функции каждого авто, еще больше запутывая будущих владельцев.
Производители прекрасно понимают, что иметь возможность одновременно эксплуатировать эти две машины несколько странно и еще более затратно. Именно поэтому большая часть оценочных суждений о том, что лучше – Мицубиси Паджеро или Тойота Прадо, опираются на мнения владельцев авто, которые в жару и мороз, в городских условиях и по бездорожью ежедневно отправляются на них в путь.
СодержаниеПоказать
Начнем по порядку
Оба автомобиля имею богатую историю. И предлагаются в четвертом поколении. Mitsubishi Pajero. Именно цифра четыре настораживает специалистов больше всего, ведь многие склоняются к версии, что последний Паджеро – лишь прилично обновленный вариант предыдущего поколения.
Эти подозрения подтверждает салон (кроме передней панели) и внешность внедорожника, которая очень напоминает модели второго поколения. Силовой агрегат 4M41 остался прежним и имеет объем 3,2 литра. Добавился новый бензиновый агрегат 6G75 объемом 3,8 литра. Напомним, что выпускается Pajero с 2006 года.
Силовой агрегат 4M41 остался прежним и имеет объем 3,2 литра. Добавился новый бензиновый агрегат 6G75 объемом 3,8 литра. Напомним, что выпускается Pajero с 2006 года.
Про раму лучше не вспоминать, потому что ее нет. Взамен ее установлена независимая подвеска, именно поэтому авто превратилось в семейное SUV-класса с расширенными возможностями полноприводной трансмиссии. Позиционировать Pajero как экстремальный внедорожник можно, но весьма осторожно.
Пассажиры и элементы ходовой части от такой конструкции страдают больше всего, особенно при езде по пересеченной местности. В этом случае подвижные части кузова подвергаются сильной нагрузке и удерживаются за счет петель, постоянно соприкасаясь с уплотнителями дверей. Со временем такой контакт приводит к протиранию резины и соприкосновению непосредственно с металлическими частями конструкции.
Особые неудобства приносит посредственная шумоизоляция. В случае дополнительной обработки антискрипом вопрос решается только на 50%.
А как обстоят дела у Prado? Land Cruiser Prado 150 вышел немного позднее – в 2009 году, но также, как и в третьем поколении, построен на той же колесной базе, что и предшественник 120, а значит, он имеет рамную конструкцию. По заявлениям представителей компании, по всей видимости, это будет последняя модификация, имеющая рамную основу.
Изменениям подверглись размеры кузова, он стал больше. В наследство от предшественника достался постоянный полный привод (в этом нет ничего плохого). Последнее поколение приобрело дополнительную опцию KDSS (в 3 поколении ее не было), осталась без изменений фирменная пневматическая подвеска Land Cruiser.
Проходимость
Один из главных критериев при выборе – проходимость. Оба автомобиля изначально были предназначены для комфортного преодоления препятствий. Сравнить Тойота Прадо и Митсубиси Паджеро можно только, если эксплуатировать их в одинаковых условиях.
В последнем авто более отзывчива трансмиссия, прежде всего, из-за грамотно проведенных заводских настроек. Электроника, обеспечивающая притормаживание колес, находящихся в воздухе, гарантирует отличное взаимодействие остальных колес с сыпучим грунтом и грязевым слоем.
Электроника, обеспечивающая притормаживание колес, находящихся в воздухе, гарантирует отличное взаимодействие остальных колес с сыпучим грунтом и грязевым слоем.
Поэтому забираться в гору на Паджеро или Прадо – большая разница. Тойота однозначно проигрывает, постоянно пробуксовывая. При нажатии на педаль акселератора в верхней части подъема наблюдается продолжительная пауза и только затем колеса начинают притормаживать. Часто этого бывает достаточно, чтобы потерять инерцию движения и загрузнуть.
В обоих авто трансмиссия находится в равном положении – включена пониженная передача с активной блокировкой межосевого дифференциала. Наличие межколесного дифференциала у Тойоты и Паджеро в бюджетных вариантах не предусмотрено. В зависимости от комплектации клиренс у обеих машин не одинаков, соответственно 220 и 235 мм. С установкой защиты он может меняться в меньшую сторону на 10-15 мм.
Нельзя не отметить, что подвеска Прадо намного комфортнее, особенно при преодолении пересеченных участков.
Интерьер
В максимальной комплектации Ultimate у Pajero IV не может не порадовать достаточно большого размера люк (его у Прадо нет), кожаный салон. Передние сидения регулируются механическим способом. Руль не регулируется по вылету.
У Land Cruiser Prado 150 в средней комплектации Elegance рулевое колесо регулируется и по высоте, и по вылету, что очень удобно, если эксплуатируется несколькими водителями. Двигатель заводится с кнопки, есть доступ без ключа.
Воспользовавшись электрическим приводом, можно установить в удобное положение передние сидушки, хотя функцию запоминания изготовители не предусмотрели. Все это можно найти в комплектации Prestige, а кроме того, присутствует навигация, блокировка межколесного дифференциала, по периметру расположены 4 камеры. Все это отображается на цене (больше на 400 тысяч).
Передняя панель на Паджеро выглядит немного изысканнее, чем у Прадо. Хотя у последней имеется цветной монитор, но с несколько архаичным разрешением 400 на 800.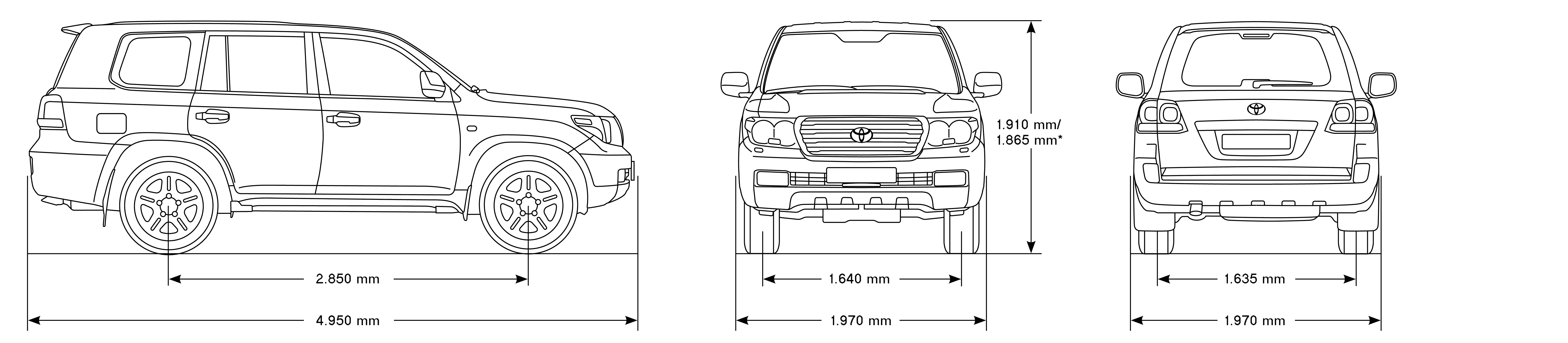 Салон предлагается только в трех вариантах: слоновая кость, черный и коричневый. Последний цвет был добавлен позже. К качеству отделки и материалам салона нет претензий по обоим авто.
Салон предлагается только в трех вариантах: слоновая кость, черный и коричневый. Последний цвет был добавлен позже. К качеству отделки и материалам салона нет претензий по обоим авто.
Отдельного внимания заслуживает вместимость салона и багажника у Паджеро – она больше. В авто есть различные ниши в задней части под полом (место под запасное колесо), куда можно поместить объемные вещи. В некоторых комплектациях авто возможно установить дополнительный ряд (машина становится 7-местной), на котором будет не очень удобно, особенно путешествуя на длинные расстояния. Но возможность разместиться все же предусмотрена.
В двух авто установлены музыкальные комбайны, в Паджеро даже высокого качества, но звук оставляет желать лучшего. Предусмотрена отличная опция с цветным монитором на потолке, это дает возможность присматривать кино и мультфильмы. Пассажирам на втором ряду будет не скучно при длительных переездах, а водителю спокойнее, так как не придется отвлекаться на разговоры, особенно если в салоне будут ехать дети.
Экономичность
Следует отметить, что в Паджеро изготовитель рекомендует заливать бензин АИ-92, а в Прадо – АИ-95. При одинаковых бензиновых агрегатах объемом 3 литра максимальная скорость у последнего на 10 км меньше и составляет 165 км/ч.
Расход на сотню у автомобилей практически одинаков и равен по трассе 10 литрам на 100 км. Поэтому Паджеро или Прадо что лучше? В этом они практически равны. А вот по городу выгоднее эксплуатировать Прадо, его расход составляет 14 литров с небольшим, против 16 с хвостиком у Паджеро.
Спорт-версия Паджеро
В 1996 году был представлен Pajero Sport. Последняя версия была презентована в 2015. Она была построена на базе Mitsubishi L200. Вопрос о том, что предпочесть – Паджеро спорт или Прадо, неоднозначен. Ведь фактически Тойота остается более мягкой по отношению даже к Pajero четвертого поколения, поэтому на комфорт в спорт-версии рассчитывать не стоит. Внедорожник приобрел абсолютно новый режим Off-Road, работающий в четырех вариантах – «гравий», снег/грязь», «песок», «камень».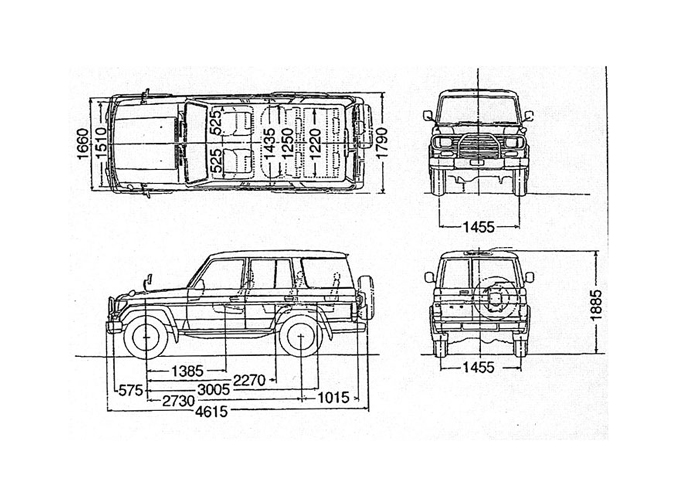 Поэтому препятствия будут преодолеваться значительно легче и безопаснее.
Поэтому препятствия будут преодолеваться значительно легче и безопаснее.
Жесткая рама стала еще жестче, а подвеска настроена на улучшенную управляемость, хотя по конструкции ее элементы остались такими же, как и в предыдущем поколении.
Что лучше Прадо или Паджеро Спорт? Учтите особенности эксплуатации. Если планируется использовать авто в агрессивном стиле, в условиях бездорожья, тогда отдайте предпочтение спорт-версии. При условии езды в городе склоняйтесь к Прадо. Не менее важным фактором при покупке является стоимость внедорожников. Цена Mitsubishi Pajero Sport дешевле, чем на Land Cruiser Prado.
А далее видео о том, что лучше Мицубиси Паджеро или Тойота Прадо:
Оцените статью
Тойота Ленд Крузер Прадо 2020-2021
Юридическая информация1 Для автомобилей в комплектациях с кожаной обивкой салона применяется комбинация из натуральной и синтетической кожи.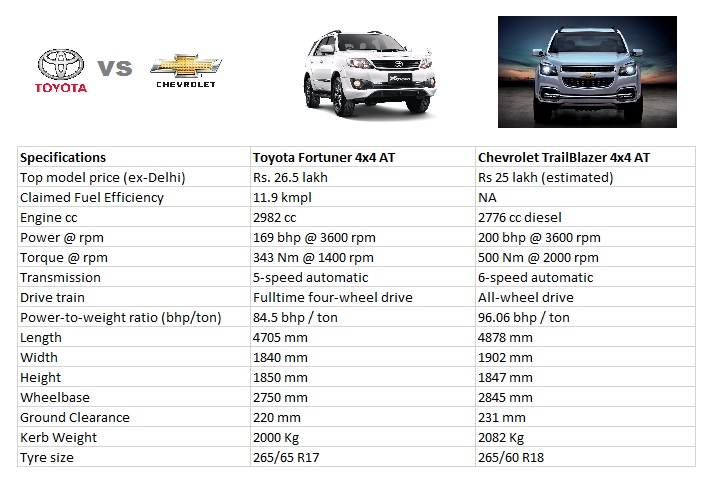
2 Набор опций может отличаться в зависимости от комплектации.
3 Новые активные системы безопасности такие, как Toyota Safety Sense, предназначены для помощи водителю. Поскольку из-за внешних факторов существует ограничение на точность распознавания и эффективность управления, обеспечиваемые данной системой, не следует слишком полагаться на систему. Водителю всегда необходимо обращать пристальное внимание на окружающую автомобиль обстановку и соблюдать все меры предосторожности при вождении. Эксплуатация Toyota Safety Sense может быть затронута или затруднена из-за внешних факторов, и Toyota не несет ответственности за какие-либо последствия, связанные с использованием системы.
4 Toyota Land Cruiser Prado признан лидером в сегменте SUV (D) (2 место в 2021 году, 1 место в 2020 году) по сохранению остаточной стоимости согласно результатам ежегодного исследования «Residual value», проведенного аналитическим агентством «АВТОСТАТ».
5 Количество автомобилей по указанным ценам ограниченно. Производитель оставляет за собой право изменять спецификацию и цены без предварительного уведомления. Информация о ценах на продукцию, модельном ряде и комплектациях носит исключительно информационный характер. Указанные цены, могут отличаться от действительных цен Уполномоченных Дилеров и/или Уполномоченных Партнеров. Приобретение любой продукции осуществляется в соответствии с условиями индивидуального договора купли-продажи. Представленная информация о продукции также не означает, что данная продукция имеется в наличии у Уполномоченных Дилеров и/или Уполномоченных Партнеров для продажи. Информацию о наличии автомобилей, а также подробные сведения об автомобилях вы можете уточнить в отделе продаж.
Производитель оставляет за собой право изменять спецификацию и цены без предварительного уведомления. Информация о ценах на продукцию, модельном ряде и комплектациях носит исключительно информационный характер. Указанные цены, могут отличаться от действительных цен Уполномоченных Дилеров и/или Уполномоченных Партнеров. Приобретение любой продукции осуществляется в соответствии с условиями индивидуального договора купли-продажи. Представленная информация о продукции также не означает, что данная продукция имеется в наличии у Уполномоченных Дилеров и/или Уполномоченных Партнеров для продажи. Информацию о наличии автомобилей, а также подробные сведения об автомобилях вы можете уточнить в отделе продаж.
6 Автомобили, представленные на фото, могут отличаться от автомобилей, доступных для заказа у официальных дилеров Тойота.
New Toyota Land Cruiser Prado 2.7 TX 7-Seater (A) Технические характеристики и характеристики Сингапур
Это прошлая / недоступная модель
В настоящее время показаны спецификации 2. 7 TX 7-Seater (A)
Объем двигателя
2693 куб. См
Конфигурация двигателя
4-цилиндровый рядный DOHC
Степень сжатия
9.6
Диаметр цилиндра x ход поршня
(95,0 x 95,0) мм
Тип топлива
Бензин
Мощность
122 кВт (163 л.с.)
Крутящий момент
246 Нм
Время разгона (0-100 км / ч)
неизвестно
Максимальная скорость
неизвестно
Расход топлива
9 км / л
Выбросы CO2
250 г / км (согласно испытаниям LTA)
Трансмиссия
6-ступенчатая (A) Super ECT
Тип привода
Полный привод
Тип
Внедорожник
Размеры (Д x Ш x В)
(4760 x 1885 x 1850) мм
Колесная база
2790 мм
Мин.
 Радиус поворота
Радиус поворота5,800 мм
Снаряженная масса
2,080 кг
Емкость топливного бака
87 л
Вместимость груза / загрузки
неизвестно
Тормоза (передние)
Дисковые вентилируемые
Тормоза (задние)
Дисковые вентилируемые
Подвеска (передняя)
Независимая двойная тяга с винтовой пружиной и стабилизаторами
Подвеска (задняя)
Продольный мост с винтовыми пружинами и стабилизаторами
 Радиус поворота
Радиус поворота| TOYOTA LAND CRUISER PRADO RX | GF-RZJ90W | 2693cc | 4AT | 3 | SUV | SUV -KZJ90W | 2982cc | 5MT | 3 | внедорожник | ||
| TOYOTA LAND CRUISER PRADO RX | KH-KZJ90W | 2982cc SUV | 9015 9015 9015 9015 9015 9015 9015 9015KH-KZJ90W | 2982cc | 4AT | 3 | внедорожник | |||||
| TOYOTA LAND CRUISER PRADO TX | KH-KZJ95W | 2982cc | RX | KH-KZJ90W | 2982cc | 5MT | 3 | внедорожник | ||||
| TOYOTA LAND CR UISER PRADO ACTIVE VACATION TX | GF-VZJ95W | 3378cc | 4AT | 5 | внедорожник | |||||||
| TOYOTA LAND CRUISER PRADO TX PACKAGE I | 9015 90 -15||||||||||||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ III | KH-KZJ95W | 2982cc | 4AT | 5 | внедорожник | |||||||
| Внедорожник | ||||||||||||
| TOYOTA LAND CRUISER PRADO TX | GF-VZJ95W | 3378cc | 4AT | 5 | SUV | |||||||
| LADISACK 9015 CR7 CR7 PRADO 9015 LADIS 9015 TUYOTACK 9014 | 4AT | 5 | внедорожник | |||||||||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TZ | GF-VZJ95W | 90 156 3378cc4AT | 5 | внедорожник | ||||||||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ I | GF-VZJ95W | 3378cc | 4AT | SUV ПАКЕТ III | GF-VZJ95W | 3378cc | 4AT | 5 | SUV | |||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX ПАКЕТ | GF-VZ6 9015 GF-VZ6 GF-VZ6 GF-VZ6||||||||||||
| TOYOTA LAND CRUISER PRADO TZ | GF-VZJ95W | 3378cc | 4AT | 5 | SUV | |||||||
| TOYOTA LAND CRUISER PRADO TZ 9015 9015 9015 9015 TX15 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 | 5 | внедорожник | ||||||||||
| TOYOTA LAND CRUISER PRADO RZ | GF-VZJ90W | 3378cc | 901 56 4AT3 | внедорожник | ||||||||
| TOYOTA LAND CRUISER PRADO АКТИВНЫЙ ОТПУСК TZ | KH-KZJ95W | 2982cc | 5MT | 9016 | KH-KZJ95W | 2982cc | 4AT | 5 | внедорожник | |||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TZ | KH-15 9015 9015AT KZJ95W | 9015 9015 9015 | CRUISER PRADO TX ПАКЕТ III | GF-VZJ95W | 3378cc | 4AT | 5 | SUV | ||||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX PACKAGE | K82 KZ157внедорожник | |||||||||||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX PACKAGE II | KH-KZJ95W | 2982 куб. см см | 4AT | 5 | внедорожник | |||||||
| TOYOTA LAND CRUISER PRADO TZ | KH-KZJ95W | 2982cc | 5MT | SUV 5 | 5MT | SU16 | KH-KZJ95W | 2982cc | 5MT | 5 | внедорожник | |
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX ПАКЕТ III | KH-KZJ95W | 9015 9015 9015 9015 9015 | TOYOTA LAND CRUISER PRADO TZ | KH-KZJ95W | 2982cc | 4AT | 5 | внедорожник | ||||
| TOYOTA LAND CRUISER PRADO PRADO ACTIVE VACATION TX7 9015 9015 | 9015внедорожник | |||||||||||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX | KH-KZJ95W | 2982cc | 4AT | 5 | внедорожник | |||||||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ III | KH-KZJ95W | 2982cc | 4AT | 5 | TOJ PRADO | KZJ 9015 | 2982cc | 4AT | 3 | SUV | ||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ II | KH-KZJ95W | 2982cc | 4AT | SUV | KH-KZJ95W | 2982cc | 5MT | 5 | внедорожник | |||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ II | KH-KZJ95W | 9015 LAND CRUISER PRADO TX ПАКЕТ III | KH-KZJ95W | 2982cc | 5MT | 5 | SUV | |||||
| TOYOTA LAND CRUISER PRADO RZ | KH-KZJ90W | 2982cc | 5MT | 3 | SUV | |||||||
| TOYOTA LAND CRUISER PRADO RZ 9015 9015 9015 9015 9015 9015 KZ7 Внедорожник | ||||||||||||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ I | KH-KZJ95W | 2982cc | 4AT | 5 | SUV | |||||||
| TXISACK PRADAGE 9015 9015 TOYEROTA PRADO LANDO 9015 5MT | 5 | внедорожник | ||||||||||
| TOYOTA LAND CRUISER PRADO RX ПАКЕТ III | KH-KZJ90W | 2982cc | 4AT | 3 | AND PRADIS | 2982cc | 4AT | 5 | SUV | |||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ II | KH-KZ J95W | 2982cc | 5MT | 5 | внедорожник | |||||||
| TOYOTA LAND CRUISER PRADO TX | KH-KZJ95W | KH-KZJ90W | 2982cc | 4AT | 3 | внедорожник | ||||||
| TOYOTA LAND CRUISER PRADO RX ПАКЕТ II | KH-KZJ90W SUV | 2982 2982 LAND CRUISER PRADO RX ПАКЕТ III | KH-KZJ90W | 2982cc | 5MT | 3 | SUV | |||||
| TOYOTA LAND CRUISER PRADO RX PACKAGE II | Внедорожник | |||||||||||
| TOYOTA LAND CRUISER PRADO TS | KH-KZJ95W | 2982cc | 4AT | 5 | 9015 6 SUV||||||||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX PACKAGE III | GF-RZJ95W | 2693cc | 4AT | 5 | TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX PACKAGE III | GF-RZJ95W | 2693cc | 4AT | 5 | SUV | ||
| SUV | ||||||||||||
| 2693cc | 4AT | 5 | внедорожник | |||||||||
| TOYOTA LAND CRUISER PRADO ACTIVE VACATION TX ПАКЕТ I | GF-RZJ95W | 2693cc | SUV АКТИВНЫЙ ОТПУСК TX | GF-RZJ95W | 2693cc | 4AT | 5 | внедорожник | ||||
| TOYOTA LAND CRUISER PRADO TX LIMITED | GF-RZJ956 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 6 | TOYOTA LAND CRUISER PRADO TX ПАКЕТ II | GF-RZJ95W | 2693cc | 4AT | 5 | внедорожник | |||||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ III | GF-RZJ95W | 2693cc | 4AT | 5 | SUV | 9015 PRADO LADIS 9015 CR7 9015 9015TOYOTISACK 9015 CR7 PRADO | TOYOTISACK 9015 | 4AT | 5 | внедорожник | ||
| TOYOTA LAND CRUISER PRADO TX ПАКЕТ III | GF-RZJ95W | 2693cc | 4AT | 9016 -RZJ95W | 2693cc | 4AT | 5 | внедорожник | ||||
| TOYOTA LAND CRUISER PRADO RX ПАКЕТ III | GF-RZJ90W | 9015AT3 SU6 2693cc6 9015ATIS 9015AT RX ПАКЕТ IIGF-RZJ90W | 2693cc | 4AT | 3 | Внедорожник | ||||||
| TOYOTA L И CRUISER PRADO TX ПАКЕТ II | GF-RZJ95W | 2693cc | 4AT | 5 | Внедорожник | |||||||
| TOYOTA LAND CRUISER PRADO RX PACKAGE I | RZJ95 9015 RZJ95 9015 Внедорожник||||||||||||
| TOYOTA LAND CRUISER PRADO TX | GF-RZJ95W | 2693cc | 4AT | 5 | SUV | |||||||
| 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 9015 | SUV |
Frontiers | Дифференциальные эффекты инактивации дискретных областей медиальной префронтальной коры на консолидацию памяти при умеренном и интенсивном обучении избеганию ингибирования
Введение
Большое количество исследований показало, что вмешательство в нейронную активность вскоре после обучения приводит к значительному дефициту консолидации памяти (McGaugh, 1966, 2000; Lechner et al. , 1999; Izquierdo and McGaugh, 2000), что сильно подтверждает гипотезу консолидации, выдвинутую Мюллером и Пилцекером (1900). Эта гипотеза предполагает, что фиксация памяти требует времени (консолидации) и что память уязвима в период консолидации. Эта гипотеза, однако, не учитывает хранение в памяти при некоторых условиях обучения.
, 1999; Izquierdo and McGaugh, 2000), что сильно подтверждает гипотезу консолидации, выдвинутую Мюллером и Пилцекером (1900). Эта гипотеза предполагает, что фиксация памяти требует времени (консолидации) и что память уязвима в период консолидации. Эта гипотеза, однако, не учитывает хранение в памяти при некоторых условиях обучения.
Было обнаружено, что изменение количества тренировок имеет важные последствия для процессов памяти. Когда обучение осуществляется посредством интенсивных тренировок, формирование памяти защищается от множества обработок амнезиаками (обзор см. В Prado-Alcalá et al., 2012). Этот защитный эффект постоянно обнаруживается после тренировки инструментальных задач, когда подкрепление доступно после выполнения определенного ответа (Prado-Alcalá и Cobos-Zapiaín, 1977, 1979; Prado-Alcalá et al., 1980). Этот эффект также был описан в задачах, которые влекут за собой как классические, так и инструментальные компоненты, такие как активное и тормозящее избегание (IA). В этих случаях животное подвергается воздействию условного раздражителя, а затем безусловного отвращающего раздражителя, независимо от его поведения.Однако после обучения животные могут избежать аверсивной стимуляции, выполняя инструментальную реакцию до появления стимула. Таким образом, вмешательство в серотонинергическую активность ухудшает как приобретение, так и сохранение активного избегания после тренировки с относительно низкой интенсивностью ударов стопы, но не при тренировке с более высокой интенсивностью ударов стопы (Galindo et al., 2008). Точно так же электролитические поражения боковых и базальных ядер миндалины затрудняли выполнение задачи избегания Сидмана, но усиленная тренировка защищала выполнение этой задачи (Lazaro-Muñoz et al., 2010).
В этих случаях животное подвергается воздействию условного раздражителя, а затем безусловного отвращающего раздражителя, независимо от его поведения.Однако после обучения животные могут избежать аверсивной стимуляции, выполняя инструментальную реакцию до появления стимула. Таким образом, вмешательство в серотонинергическую активность ухудшает как приобретение, так и сохранение активного избегания после тренировки с относительно низкой интенсивностью ударов стопы, но не при тренировке с более высокой интенсивностью ударов стопы (Galindo et al., 2008). Точно так же электролитические поражения боковых и базальных ядер миндалины затрудняли выполнение задачи избегания Сидмана, но усиленная тренировка защищала выполнение этой задачи (Lazaro-Muñoz et al., 2010).
Системное лечение амнезиаками также препятствует консолидации памяти IA, но, опять же, такого дефицита не возникает после интенсивных тренировок (Durán-Arévalo et al., 1990; Cruz-Morales et al., 1992; Solana-Figueroa et al., 2002; Диас-Трухильо и др. , 2009). То же самое верно и при лечении полосатого тела (Giordano and Prado-Alcalá, 1986; Pérez-Ruiz and Prado-Alcalá, 1989; Salado-Castillo et al., 2011), гиппокампа (Quiroz et al., 2003; Garín -Aguilar et al., 2014), миндалевидное тело (Parent et al., 1992, 1994; Тэтчер и Кимбл, 1966) и черной субстанции (Cobos-Zapiaín et al., 1996).
, 2009). То же самое верно и при лечении полосатого тела (Giordano and Prado-Alcalá, 1986; Pérez-Ruiz and Prado-Alcalá, 1989; Salado-Castillo et al., 2011), гиппокампа (Quiroz et al., 2003; Garín -Aguilar et al., 2014), миндалевидное тело (Parent et al., 1992, 1994; Тэтчер и Кимбл, 1966) и черной субстанции (Cobos-Zapiaín et al., 1996).
Медиальная префронтальная кора (mPFC), которая включает переднюю поясную извилину (ACC), прелимбическую (PL) и инфралимбическую (IL) области (Heidbreder and Groenewegen, 2003; Vertes, 2004, 2006), получила большое внимание. в связи с его участием в классической условной рефлексии страха (Mello e Souza et al., 1999; Yang and Liang, 2014; Zhang et al., 2011), и было высказано предположение, что эти области участвуют по-разному на разных этапах запоминания обусловливание страхом (Джустино и Марен, 2015).Таким образом, ACC был связан с приобретением (Sacchetti et al., 2003; Tang et al., 2005; Bissière et al., 2008), PL — с экспрессией (извлечением) (Blum et al. , 2006; Vidal-Gonzalez et al. ., 2006; Corcoran, Quirk, 2007) и IL с процессом вымирания (Quirk, Mueller, 2008) и контролем над страхом (Sotres-Bayon and Quirk, 2010). Однако литература об участии mPFC в процессах памяти, связанных с инструментальным исполнением, немногочисленна. Сообщалось, что электролитические поражения IL, но не PL, вызывают дефицит инструментального компонента, участвующего в сохранении пониженного IA (Jinks and McGregor, 1997).
, 2006; Vidal-Gonzalez et al. ., 2006; Corcoran, Quirk, 2007) и IL с процессом вымирания (Quirk, Mueller, 2008) и контролем над страхом (Sotres-Bayon and Quirk, 2010). Однако литература об участии mPFC в процессах памяти, связанных с инструментальным исполнением, немногочисленна. Сообщалось, что электролитические поражения IL, но не PL, вызывают дефицит инструментального компонента, участвующего в сохранении пониженного IA (Jinks and McGregor, 1997).
Насколько нам известно, защитный эффект усиленного обучения на обучение и память не изучался в отношении избирательной инактивации нейронной активности ACC, PL и IL. По этой причине, а также из-за различных функциональных атрибутов, описанных в mPFC, мы сочли важным изучить эффекты временной инактивации трех областей mPFC на консолидацию памяти при умеренной и интенсивной тренировке IA. Мы предположили, что временная инактивация AC, PL и IL будет оказывать различное влияние на консолидацию памяти IA, и что интенсивное обучение компенсирует потенциальные недостатки, вызванные такой инактивацией.
Материалы и методы
В этом разделе описаны процедуры, общие для всех экспериментов данного исследования. При необходимости будут описаны другие процедуры, характерные для конкретных экспериментов.
Субъекты
крыс-самцов линии Wistar (300–350 г) из колонии для разведения в Instituto de Neurobiología, Universidad Nacional Autónoma de México, содержали индивидуально с водой и пищей ad libitum и содержали в комнате со светом 12 ч / 12 ч. -темный цикл (свет включается в 7:00).Температура в помещении составляла 23 ± 1 ° C. Крыс случайным образом распределяли по каждой группе, а обучение и тестирование проводили во время легкой фазы цикла с 8:00 до 12:00. Протокол эксперимента был одобрен Комитетом по этике животных Института нейробиологии, Национальный автономный университет Мексики и соответствовал Руководству по уходу и использованию лабораторных животных (Комитет Национального исследовательского совета (США) по обновлению Руководства по уходу и Использование лабораторных животных, 2011 г. ).
).
Хирургия
Крыс анестезировали пентобарбиталом натрия (50 мг / кг, внутрибрюшинно), вводили атропин (1 мг / кг, внутрибрюшинно) для предотвращения обструкции дыхательных путей, и их головы помещали на стереотаксическую рамку (Stoelting Co., United Состояния). Наконечники двусторонней направляющей канюли из нержавеющей стали (длина: 8 мм для ACC и 10 мм для PL и IL; калибр 23) были нацелены на 1 мм выше ACC (+2,8 мм от брегмы; ± 0,4 мм от средней линии; -1,4 мм. ниже черепа), PL (+3,0 мм от брегмы; ± 0.7 мм от средней линии; -3,2 мм ниже черепа) или IL (+3,0 мм от брегмы; ± 0,6 мм от средней линии; -4,2 мм ниже поверхности черепа) (Paxinos and Watson, 2007). Канюли прикрепляли к черепу с помощью одного ювелирного винта и стоматологического цемента. Стилеты (длиной 8 мм для ACC и длиной 10 мм для PL и IL) вставляли в каждую канюлю для поддержания проходимости, а затем удаляли и помещали обратно во время сеансов манипуляции и для введения лечения. После операции животные получали 1,0 мл 0,9% физиологического раствора, внутрибрюшинно, и находились в инкубаторе до полного выхода из наркоза. После операции крысам давали возможность восстановиться в течение 7 дней перед началом обучения. В течение этого периода экспериментатор прикасался к каждому животному, осторожно касаясь и удерживая крысу в течение приблизительно 5 минут в течение трех дней подряд.
После операции крысам давали возможность восстановиться в течение 7 дней перед началом обучения. В течение этого периода экспериментатор прикасался к каждому животному, осторожно касаясь и удерживая крысу в течение приблизительно 5 минут в течение трех дней подряд.
Аппарат
Крысы обучались в аппарате ИА, состоящем из двух отсеков, разделенных раздвижной дверью. Сейф (30 см × 30 см × 30 см) имел крышку и стенки из прозрачного акрила красного цвета, а дно — из стержней из нержавеющей стали (диаметром 6 мм, 9 мм друг от друга).Этот отсек освещался лампочкой мощностью 10 Вт, расположенной в центре его крышки. Другой, неосвещенный противоударный отсек (длина 30 см) имел переднюю и заднюю стенки и пол из пластин из нержавеющей стали с боковыми стенками и крышкой из прозрачного акрила красного цвета. Стены и пол имели форму желоба, шириной 20 см вверху и 8 см внизу. В середине пола прорезь 1,5 см разделяла две пластины из нержавеющей стали, из которых состоят стены и пол. При входе в неосвещенный отсек крысы контактировали с обеими пластинами, через которые можно было произвести удар ногой. Стимулятор прямоугольных импульсов (модель Grass S-48), подключенный последовательно к устройству постоянного тока (модель Grass CCU-1), генерировал толчок стопы. Доставка удара и измерение времени ожидания перехода из одного отсека в другой выполнялись с помощью автоматизированного оборудования. Оба отсека протирали 10% спиртом до и после того, как каждая крыса заняла его. Аппарат находился в темном звукоизолированном помещении с фоновым маскирующим шумом.
Стимулятор прямоугольных импульсов (модель Grass S-48), подключенный последовательно к устройству постоянного тока (модель Grass CCU-1), генерировал толчок стопы. Доставка удара и измерение времени ожидания перехода из одного отсека в другой выполнялись с помощью автоматизированного оборудования. Оба отсека протирали 10% спиртом до и после того, как каждая крыса заняла его. Аппарат находился в темном звукоизолированном помещении с фоновым маскирующим шумом.
Обучение и тестирование избегания ингибирования
В день обучения каждую крысу помещали в безопасный отсек, а через 10 с открывали дверь между двумя отсеками.Задержка перехода от безопасного отсека к шоковому отсеку называется задержкой обучения. Как только животные переходили в это отделение, дверь закрывалась, и производился удар ногой 1,0 или 3,0 мА (последовательность прямоугольных импульсов 50 мс с частотой 10 Гц). Через пять секунд дверь снова открыли, позволив крысе сбежать в безопасный отсек, а затем выключили стимулятор; эта задержка называется задержкой выхода. Через 30 с в безопасном отсеке крысу помещали обратно в домашнюю клетку.Удержание задания измерялось через 48 часов после тренировки; в некоторых случаях удерживание регистрировали как через 30 мин (во время получения кодирования), так и через 48 ч после обучения у одних и тех же животных. В этих сеансах удержания применялась та же процедура, что и на тренировке, за исключением того, что удары ногой не применялись. Если крыса не пересекалась в течение 600 с, сеанс заканчивался и присваивалась оценка 600.
Через 30 с в безопасном отсеке крысу помещали обратно в домашнюю клетку.Удержание задания измерялось через 48 часов после тренировки; в некоторых случаях удерживание регистрировали как через 30 мин (во время получения кодирования), так и через 48 ч после обучения у одних и тех же животных. В этих сеансах удержания применялась та же процедура, что и на тренировке, за исключением того, что удары ногой не применялись. Если крыса не пересекалась в течение 600 с, сеанс заканчивался и присваивалась оценка 600.
Процедуры
Тетродотоксин (TTX) был использован для инактивации целевых областей; он обратимо блокирует зависимые от напряжения натриевые каналы, тем самым предотвращая генерацию и распространение потенциалов действия (Fozzard and Lipkind, 2010).Одновременные двусторонние инфузии ТТХ (Sigma, C11h27N3O8, T8024; 0,3 мкг / полушарие, растворенные в 0,3 мкл изотонического раствора) или равного объема носителя (VEH) в ACC, PL или IL проводились за 25 минут до тренировки. В дополнительных группах крыс ту же дозу ТТХ или VEH вводили в ИЛ сразу после обучения. Скорость инфузии составляла 0,3 мкл / мин и контролировалась автоматическим микроинфузионным насосом (WPI, модель 220i). По окончании инфузии иглы для инъекций, которые выступали 1.0 мм за кончиком канюли, оставалось внутри направляющей канюли в течение 60 с, чтобы минимизировать обратный ток. Процедура инъекции проводилась в другом помещении, чем то, в котором проходили обучение и тестирование.
В дополнительных группах крыс ту же дозу ТТХ или VEH вводили в ИЛ сразу после обучения. Скорость инфузии составляла 0,3 мкл / мин и контролировалась автоматическим микроинфузионным насосом (WPI, модель 220i). По окончании инфузии иглы для инъекций, которые выступали 1.0 мм за кончиком канюли, оставалось внутри направляющей канюли в течение 60 с, чтобы минимизировать обратный ток. Процедура инъекции проводилась в другом помещении, чем то, в котором проходили обучение и тестирование.
Гистология
Крыс анестезировали пентобарбиталом натрия (125 мг / кг) и внутрисердечно перфузировали 0,9% физиологическим раствором с последующим введением 4% формалина. Мозги извлекали и погружали в 4% раствор формальдегида минимум на 5 дней. Срезы (толщиной 50 мкм) вырезали на криостате и окрашивали крезиловым фиолетовым.Срезы исследовали под световым микроскопом и определяли расположение кончиков инъекционных игл. Данные о крысах с кончиками канюли за пределами целевых областей не были включены в статистический анализ. На рисунках 1C, 2F и 3F показаны примеры участков кончика канюли в ACC, PL и IL соответственно.
На рисунках 1C, 2F и 3F показаны примеры участков кончика канюли в ACC, PL и IL соответственно.
РИСУНОК 1. Передняя поясная корка. (A) Средний балл удержания (с межквартильным размахом) групп крыс, обученных в одном испытании IA и которым за 25 минут до обучения вводили раствор носителя (VEH) или TTX в переднюю поясную извилину коры головного мозга (ACC).TTX был неэффективен в изменении консолидации памяти при умеренной (1,0 мА) или интенсивной (3,0 мА) тренировке. (B) TTX вызывает снижение экспрессии c-Fos вблизи кончика инжектора. ∗ p <0,05 по сравнению с VEH. (C) Типичная микрофотография, показывающая расположение канюли (толстая стрелка) и кончиков инъекционных игл (тонкая стрелка). Числа внутри столбцов представляют размер выборки.
РИСУНОК 2. Прелимбическая кора. Медианные оценки удержания (с межквартильными диапазонами) групп крыс, обученных в одном испытании IA и которым за 25 минут до тренировки вводили VEH или TTX в прелимбическую кору (PL). (A) TTX нарушение консолидации памяти средней (1,0 мА), но не интенсивной (3,0 мА) тренировки. (B) TTX не мешает приобретению (ACQ), но значительно ухудшает долговременную память (LTM). (C) TTX не создавал зависимости от состояния; ТТХ и VEH вводили дважды, за 25 мин до тренировки и за 25 мин до теста на удержание. Дефицит удержания наблюдался только после лечения ТТХ. (D) TTX вызывает снижение экспрессии c-Fos вблизи кончика инжектора. ∗ p <0,05 относительно VEH. (E) TTX не изменял экспрессию c-Fos за пределами 600 мкм ниже кончика канюли. (F) Типичная микрофотография, показывающая расположение канюли (толстая стрелка) и кончиков инъекционных игл (тонкая стрелка). Числа внутри столбцов представляют размер выборки.
(A) TTX нарушение консолидации памяти средней (1,0 мА), но не интенсивной (3,0 мА) тренировки. (B) TTX не мешает приобретению (ACQ), но значительно ухудшает долговременную память (LTM). (C) TTX не создавал зависимости от состояния; ТТХ и VEH вводили дважды, за 25 мин до тренировки и за 25 мин до теста на удержание. Дефицит удержания наблюдался только после лечения ТТХ. (D) TTX вызывает снижение экспрессии c-Fos вблизи кончика инжектора. ∗ p <0,05 относительно VEH. (E) TTX не изменял экспрессию c-Fos за пределами 600 мкм ниже кончика канюли. (F) Типичная микрофотография, показывающая расположение канюли (толстая стрелка) и кончиков инъекционных игл (тонкая стрелка). Числа внутри столбцов представляют размер выборки.
РИСУНОК 3. Инфралимбическая кора. Медианные оценки удержания (с интерквартильными диапазонами) групп крыс, обученных в одном испытании IA и которым за 25 минут до тренировки вводили VEH или TTX в инфралимбическую кору (IL). (A) TTX нарушает удержание умеренной (1,0 мА) и интенсивной (3,0 мА) тренировки. (B) TTX не препятствовал получению, но значительно ухудшал LTM. (C) TTX и VEH вводили дважды, за 25 минут до тренировки и за 25 минут до теста на удержание. Дефицит удержания наблюдался в группе ТТХ с умеренным тренингом, а зависимость от состояния возникла в группе ТТХ с интенсивным обучением, которая показала хорошее удержание задания. (D) Введение ТТХ сразу после тренировки вызывало дефицит консолидации памяти после умеренной тренировки (1.0 мА), но не после интенсивной тренировки (3,0 мА). (E) TTX вызывает снижение экспрессии c-Fos вблизи кончика инжектора. ∗ p <0,05 относительно VEH. (F) Типичная микрофотография, показывающая расположение канюли (толстая стрелка) и кончиков инъекционных игл (тонкая стрелка). Числа внутри столбцов представляют размер выборки.
c-FOS Иммуногистохимия
Чтобы оценить блокирующий эффект ТТХ на нервную активность в каждой из трех исследованных областей mPFC, мы использовали иммуногистохимию для обнаружения c-Fos, поскольку этот белок обычно используется в качестве маркера такой активности (Sagar et al., 1988; Эррера и Робертсон, 1996; Уиллоуби и др., 1997). С этой целью для каждой области группа крыс обучалась с 1,0 или 3,0 мА, а половина группы получала TTX или VEH, как описано выше, но удержание задачи не измерялось. Группа наивных животных ( n = 6), использованная для получения базального количества c-Fos-положительных клеток, содержалась в тех же условиях жизни, что и остальные группы, но они никогда не покидали биотерию, за исключением для жертвоприношения. Через час после обучения животных анестезировали пентобарбиталом натрия (125 мг / кг) и транскардиально перфузировали физиологическим раствором с последующим введением 4% PFA (pH 9.5, 10 ° С). Мозги удаляли и хранили в фиксирующем растворе в течение 4 ч, затем в 15% сахарозе в течение ночи, а затем в 30% сахарозе; растворы хранили при 4 ° C. Через три дня четыре серийных коронарных среза (толщиной 30 мкм) были получены при -20 ° C из ACC, PL и IL и сохранены в растворе криопротектора (30% этиленгликоля и 20% глицерина в 0,05 М натрий-фосфатном буфере). при -20 ° C до гистохимической обработки. Срезы головного мозга последовательно инкубировали в 0,1 М PB в течение 20 мин, H 2 O 2 0.03% в течение 10 минут, NaBH 4 1% в течение 6 минут и NGS 3% в течение 30 минут. Затем их инкубировали в течение 48 часов при 4 ° C в поликлональных антителах c-Fos (Anti-c-Fos rabbit, 1: 5000, Abcam, Кембридж, Массачусетс, США) с последующей инкубацией в течение 1 часа в козьих антителах против кроликов. биотинилированное вторичное антитело (ВА-1000, 1: 500; Vector Laboratories, Burlingame, CA, США), 1 час в наборе Vectastain ABC Kit (Vector Laboratories, Burlingame, CA, США) и 10 мин в растворе DAB (0,03 % H 2 O 2 , NAS).Срезы головного мозга помещали на предметные стекла, постепенно обезвоживали спиртом, затем очищающим агентом ксилолом, а затем покрывали Entellan ® .
Цифровые изображения были получены с помощью Leica AF6000 Microsystem (Leica, Германия) с использованием объектива 10x. Подсчет c-Fos-положительных клеток производился автоматически с помощью плагина «счетчик клеток» с использованием программного обеспечения ImageJ (NIH). Три счетных бокса (100 мкм × 100 мкм) были расположены горизонтально и центрированы на 100 мкм ниже дорожек двусторонней инъекции иглы; таким образом, было проанализировано шесть изображений на животное.Поскольку ACC, PL и IL находятся рядом друг с другом вдоль дорсально-вентрального измерения, было важно оценить возможность того, что TTX мог иметь эффект из-за диффузии из целевой области в ее соседнюю вентральную область. С этой целью счетный ящик был расположен на 600 мкм ниже каждой дорожки кончика инъекционной иглы в группе PL. Уровень экспрессии c-Fos в каждой области мозга для каждой группы выражали как отношение среднего количества c-Fos-положительных клеток каждой крысы для каждой группы к среднему количеству c-Fos-положительных клеток для соответствующей наивная группа.
Статистический анализ
Поскольку измерение удержания задачи IA было усечено на 600 с, непараметрическая статистика использовалась при анализе поведенческих результатов. Сравнение латентности обучения, ухода и задержки между группами TTX и VEH в каждой области mPFC проводили с использованием теста Mann-Whitney U . Аналогичным образом, количество c-Fos-положительных клеток в группах TTX и VEH в каждой области mPFC сравнивали с тестом Mann-Whitney U .
Результаты
Передняя поясная корка
Задержки при обучении и побеге
Тест Mann – Whitney U показал, что не было значительных различий в показателях латентности между группами TTX и VEH, независимо от интенсивности ударов стопы, которые использовались во время тренировки. Медианные задержки тренировки в группах VEH и TTX, которые тренировались с 1,0 мА, составляли 15,85 и 30,35 с ( p = 0,15), а для тех, кто тренировался с 3,0 мА, составляли 19.70 и 31,05 с ( p = 1,0) соответственно (данные не показаны). Точно так же не было значительных различий в латентности побега между группами TTX и VEH, независимо от интенсивности шока стопы. Медианная латентность ухода, отображаемая группами 1,0 мА, составляла 4,10 и 2,30 с ( p = 0,57) соответственно. В группах VEH и TTX, обученных с 3,0 мА, латентность ухода составляла 1,35 и 1,90 с ( p = 0,68), соответственно (данные не показаны).
Долговременная память
Не было очевидных различий между группами VEH и TTX в латентности удерживания, измеренной через 48 часов после тренировки, независимо от интенсивности толчка стопы, используемого для тренировки (1.0 мА, p = 0,46 и 3,0 мА, p = 0,81) (рисунок 1A).
c-Fos Иммуногистохимия
Введение тетродотоксина в ACC вызывало значительное снижение экспрессии c-Fos по сравнению с VEH в группах, которые тренировались с 1,0 и 3,0 мА ( p <0,05 для каждой интенсивности) (рис. 1B). Рисунок 1C представляет собой типичную микрофотографию, показывающую размещение наконечника канюли в ACC.
Прелимбическая кора
Задержки при обучении и побеге
Тест Манна – Уитни U показал, что не было значительных различий в латентности обучения, отображаемой 1.0 мА TTX (40,20 с) и VEH (26,20 с) группы ( p = 0,18), а также в группах 3,0 мА TTX (28,80 с) и VEH (26,40 с) ( p = 1,0). Точно так же не было значительных различий в латентности побега между группами TTX и VEH, независимо от интенсивности шока стопы. Медианная латентность ухода, отображаемая группами 1,0 мА, составляла 1,80 и 1,40 с, соответственно ( p = 0,49), а медианная латентность ухода в группах TTX и VEH, обученных с 3,0 мА, составляла 0,80 и 0.60 с соответственно ( p = 0,27) (данные не показаны).
Долговременная память
Группа TTX, обученная с помощью 1,0 мА, показала значительно более низкий балл, чем ее контрольная группа VEH ( p <0,05) во время 48-часового теста удержания. Напротив, никаких различий не было обнаружено при сравнении групп TTX и VEH, тренированных с 3,0 мА ( p = 0,13) (рис. 2A).
Приобретение
Оценить, насколько амнестический эффект предтренировочного вливания ТТХ в 1.Группа 0 мА PL была связана с вмешательством в обучение задаче IA, а не с консолидацией, TTX или VEH вводили за 25 минут до тренировки с 1,0 мА, а удерживание измеряли дважды: через 30 минут и через 48 часов после тренировки. Результаты не показали значительных различий между группами TTX и VEH в тесте удержания через 30 минут после тренировки ( p = 0,95), в то время как, опять же, достоверный дефицит был показан группой TTX в 48-часовом тесте ( p <0,05) (Рисунок 2B).
Государственное подчинение
Поскольку ТТХ вводили за 25 минут до тренировки, а удерживание было измерено через 48 часов в немедикаментозном состоянии, было возможно, что вызванная таким образом амнезия могла быть вызвана зависимым от состояния эффектом. Чтобы исключить эту возможность, две группы крыс обрабатывались дважды, либо TTX, либо VEH, за 25 минут до обучения и за 25 минут до тестирования удержания. По сравнению с группой VEH, группа TTX показала надежную амнезию ( p <0.05) (Рисунок 2C).
c-Fos Иммуногистохимия
Введение тетродотоксина в PL вызвало значительное снижение экспрессии c-Fos по сравнению с VEH в группах, которые тренировались с 1,0 или 3,0 мА ( p <0,05, в каждом случае) (рис. 2D).
Как упоминалось в разделе «Материалы и методы», экспрессия c-Fos также измерялась в счетной ячейке 100 мкм × 100 мкм, расположенной на 600 мкм ниже дорожек кончика PL инжектора. Мы обнаружили, что TTX не влиял на экспрессию c-Fos, поскольку не было значительных различий между группами VEH и TTX, p = 0.73 (Рисунок 2E). Это открытие демонстрирует, что дефицит консолидации памяти, наблюдаемый у животных, которых тренировали с 1,0 мА, был вызван инактивацией PL, а не диффузией лекарства в более вентрально расположенный IL.
Рисунок 2F представляет собой типичную микрофотографию, показывающую размещение кончика канюли в PL.
Инфралимбическая кора
Задержки при обучении и побеге
Тест Mann – Whitney U показал, что не было значительных различий в тренировочной латентности между группами TTX и VEH, независимо от интенсивности удара стопы.Медианные задержки тренировки, отображаемые группами 1,0 мА, составили 33,60 и 25,20 с ( p = 0,80) соответственно. В группах VEH и TTX, тренированных с 3,0 мА, латентность обучения составляла 24,20 и 23,50 с ( p = 0,97) соответственно (данные не показаны). Точно так же не было значительных различий в латентности побега между группами TTX и VEH, независимо от интенсивности шока стопы. Медианная латентность ухода, отображаемая группами 1,0 мА, составляла 1,70 и 1,20 с ( p = 0.28) соответственно. В группах VEH и TTX, обученных с 3,0 мА, латентность ухода составляла 2,70 и 2,30 с ( p = 0,65), соответственно (данные не показаны).
Как и в случае PL, инфузия TTX в IL вызвала значительный ретенционный дефицит в течение 48 часов после тренировки в группе, которая тренировалась с 1,0 мА ( p <0,01 по сравнению с VEH). Неожиданно ТТХ произвел такой же амнестический эффект в группе 3,0 мА ( p <0,005 по сравнению с VEH) (рис. 3A).
Приобретение
Чтобы оценить, был ли амнестический эффект предтренировочной инфузии ТТХ обусловлен вмешательством в обучение, а не с консолидацией, две группы крыс были обучены с низким толчком стопы (1,0 мА) и подвергнуты инъекциям ТТХ или VEH в ИЖ за 25 мин до тренировки. Удержание измеряли дважды: через 30 мин и через 48 ч после тренировки. Результаты не показали значительных различий между группами TTX и VEH во время пробного запуска через 30 минут после тренировки ( p = 0.85), в то время как достоверный дефицит был показан группой ТТХ в 48-часовом тесте ( p <0,02) (Рисунок 3B).
Государственное подчинение
Чтобы определить, может ли амнестический эффект предтренировочного ТТХ в группах 1,0 и 3,0 мА IL во время 48-часового теста удерживания (рис. 3A) быть обусловлен зависимостью от состояния, две группы крыс обучались с использованием 1,0 мА. и еще две группы были обучены с 3,0 мА. Половина каждой группы получала дважды TTX, а другая половина — VEH, также дважды, за 25 минут до тренировки и за 25 минут до тестирования удержания.ТТХ давал значительно более низкий показатель удержания по сравнению с контрольной группой VEH после тренировки с группой 1,0 мА ( p <0,05). Напротив, зависимость от состояния создавалась, когда для обучения использовалось 3,0 мА, поскольку не было достоверных различий в показателях удержания между группами TTX и VEH, которые тренировались с 3,0 мА ( p = 0,11) (рис. 3C).
Введение TTX после обучения в IL
Результаты предыдущего эксперимента показали, что предтренировочная инфузия ТТХ в ИЖ давала явный зависимый от состояния эффект, когда тренировка проводилась с 3.0 мА, но не при использовании 1,0 мА. Таким образом, очевидный амнестический эффект, возникающий после однократной предтренировочной инфузии ТТХ (рис. 3A), можно объяснить взаимодействием сильного удара стопы и дифференциального фармакологического состояния коры IL во время тренировки (состояние наркотического опьянения) и тестирования удержания ( состояние без наркотиков). Этот результат не позволил нам ответить на вопрос, играет ли IL роль в консолидации памяти, когда для обучения используется сильная аверсивная стимуляция. Чтобы пролить свет на этот вопрос, мы решили изучить эффекты инактивации IL с помощью TTX, индуцированной после обучения, таким образом избегая смешивающего эффекта зависимости от состояния.С этой целью две группы крыс были обучены с использованием 1,0 мА, а еще две группы были обучены с помощью 3,0 мА. Половина каждой группы получала TTX, а другая половина — VEH. Настои готовили сразу после тренировки. Значительный дефицит удерживания наблюдался в группе TTX, которая тренировалась с 1,0 мА ( p <0,03 по сравнению с VEH), тогда как не было обнаружено значительных различий между группами TTX и VEH, тренированными с 3,0 мА ( p = 0,11 ) (Рисунок 3D).
Иммуногистохимия c-FOS
Вливание TTX в IL вызывало значительное снижение экспрессии c-Fos по сравнению с VEH в группах, которые тренировались с 1.0 или 3,0 мА ( p <0,05 для каждого сравнения) (рис. 3E).
Обсуждение
Основные результаты этого исследования, в котором TTX вводили перед тренировкой, заключались в том, что независимо от интенсивности тренировки временная инактивация ACC не нарушала консолидацию памяти при выполнении задания IA. Напротив, в PL и IL TTX вызывал очень значительный дефицит консолидации, когда при тренировке использовался умеренный толчок стопы. Интересно, что дефицит удерживания все еще был очевиден после тренировки с сильным ударом стопы, когда IL была инактивирована, из-за зависимости от состояния, но удержание не уменьшилось в группе PL (Рисунки 1A, 2A, 3A).Когда TTX вводился сразу после тренировки в IL, он мешал консолидации только при использовании умеренного толчка стопы. Мы предполагаем, что эти дифференциальные эффекты зависят от разнородной связности трех исследуемых регионов. Они получают прочные связи из одних и тех же таламических областей; PL и IL получают афференты из базолатерального и базомедиального ядер миндалины; и PL более плотно связан с лимбическими областями коры, чем ACC и IL (Hoover and Vertes, 2007).Необходимы дальнейшие исследования, чтобы изучить вклад этих различных анатомических взаимодействий в консолидацию памяти IA.
Гистохимические результаты показали, что введение ТТХ в каждую из этих областей коры приводит к надежной инактивации нейронов, о чем свидетельствует уменьшение обнаружения c-Fos возле кончиков инъекторов. Поскольку не было значительных различий в обучении и латентности ухода между животными, получавшими TTX и VEH, независимо от области микроинъекции или интенсивности толчка стопы, используемого для обучения, нарушение удержания, которое было обнаружено в группах PL и IL. не может быть объяснено каким-либо потенциальным недостатком двигательной или перцептивной активности, необходимой для выполнения задачи ВА.Другими словами, животные, подвергшиеся лечению, могли перейти из безопасного отсека в отсек шока и уйти от удара ногой так же эффективно, как и животные, леченные VEH. Далее следует обсуждение соответствующих исследований по IA.
Передняя поясная корка
Инактивация ACC не мешала консолидации памяти IA, на что указывают высокие показатели удержания у животных, обученных с 1,0 и 3,0 мА. Это согласуется с отчетом Mello e Souza et al.(1999), что введение мусцимола и AP5 внутри АСС не препятствовало формированию долговременной памяти об этой задаче. Наш результат также согласуется с отсутствием нарушения консолидации памяти IA, обнаруженным после радиочастотного поражения перед тренировкой (Chai et al., 2010) ACC. Взятые вместе, эти данные предполагают, что эта область не участвует в кодировании нейронной активности, необходимой для консолидации памяти ассоциации CS-UCS во время обучения задаче IA. К этой интерпретации следует относиться с осторожностью, потому что другие направления исследований предполагают, что ACC участвует в консолидации памяти IA.Таким образом, инфузия холинергического агониста оксотреморина в ACC сразу после тренировки IA улучшила память (Malin and McGaugh, 2006), и, в соответствии с этим открытием, было показано, что инфузия скополамина, холинергического антагониста, до и после тренировки, нарушала консолидация памяти об этой задаче (Riekkinen et al., 1995). Кроме того, введение ингибитора синтеза белка в ACC или mPFC (включая PL и IL) вызывает значительный дефицит удержания IA (Zhang et al., 2011). Чтобы понять эти несходные результаты, необходимы новые исследования.
Прелимбическая кора
Выводы о том, что инактивация PL вызвала заметный дефицит удержания, когда для тренировки использовался удар стопой низкой интенсивности, и что это не препятствовало работе при использовании сильного удара стопой (рис. 2A), хорошо согласуются с предыдущими результатами, в которых вмешательство в нервную активность полосатого тела (Giordano, Prado-Alcalá, 1986; Pérez-Ruiz and Prado-Alcalá, 1989), гиппокампа (Quiroz et al., 2003; Garín-Aguilar et al., 2014), миндалевидное тело (Parent et al., 1992, 1994) и черная субстанция (Cobos-Zapiaín et al., 1996; Salado-Castillo et al., 2011) нарушали консолидацию памяти при тренировках IA. имело место при низкой интенсивности аверсивной стимуляции, но не при использовании стимуляции относительно высокой интенсивности.
То, что нарушение удержания, показанное группой TTX, которая тренировалась с низким толчком стопы, было связано с вмешательством в консолидацию памяти, а не с недостатком обучения, было продемонстрировано оптимальной производительностью, показанной группой животных, которые был протестирован через 30 мин после введения препарата.Дефицит консолидации стал очевидным, когда этой группе был проведен второй тест на удерживание через 48 часов (рис. 2В).
Поскольку тренировка проходила под влиянием ТТХ, а удержание задания измерялось, когда животные находились в состоянии без наркотиков, существовала возможность того, что дефицит удержания, наблюдаемый в группе с низким ударом стопы, был вызван феноменом государственная зависимость, а не нарушение консолидации памяти. Эта возможность была отвергнута, потому что группа крыс, которые были обучены и протестированы в одинаковых фармакологических условиях, показала недостаточное удерживание (рис. 2С).
Дефицит удержания, наблюдаемый в настоящем исследовании, подтверждает выводы Сантос-Андерсона и Руттенберга (1976) и Джинкса и МакГрегора (1997). Первые авторы показали, что электрическая стимуляция низкого уровня вентральной части mPFC мешает консолидации памяти IA, а вторые обнаружили, что электролитические поражения PL вызывают дефицит IA. В совокупности эти данные указывают на то, что PL имеет схожие функции с функциями полосатого тела, гиппокампа, миндалины и черной субстанции в отношении консолидации памяти IA, т.е.е., эти структуры необходимы для консолидации памяти в условиях умеренного обучения, поскольку вмешательство в нейронную активность любой из них препятствует формированию долговременной памяти. С другой стороны, консолидация происходит после интенсивных тренировок, несмотря на это вмешательство. Была выдвинута гипотеза, что эти структуры не критичны для посредничества ассоциативных процессов, происходящих из интенсивной тренировки, которая производит пластические изменения, позволяющие задействовать другие структуры.
Инфралимбическая кора
Исследование IL дало сложный набор данных. В соответствии с предыдущими результатами, когда поражения IL приводили к значительному дефициту удержания IA (Jinks and McGregor, 1997), мы обнаружили, что введение ТТХ в IL имело такой же пагубный эффект, когда для тренировки использовался низкий толчок стопы. Вопреки нашим ожиданиям, интенсивные тренировки не защитили консолидацию памяти от инактивации, вызванной TTX (рис. 3A). Тест памяти, который был проведен через 30 минут после тренировки для группы животных, которым вводили ТТХ в ИЛ, дал наивысшие оценки удержания, в то время как достоверный дефицит наблюдался через 48 часов в этой же группе.Как и в случае инактивации PL, эти результаты указывают на то, что обучение имело место и что нарушение удержания было связано с неудачей в консолидации памяти (рис. 3B).
Когда ТТХ был введен в ИЛ дважды (как перед тренировкой, так и перед 48-часовым тестом на удержание), дефицит удерживания возник в группе с низким толчком стопы, но неожиданно не в группе с высоким толчком стопы (рис. 3С). Насколько нам известно, это первый случай, когда эффект зависимости от состояния был обнаружен в mPFC.Таким образом, вопрос о том, участвует ли IL в консолидации памяти при интенсивных тренировках, не может быть решен с помощью этого экспериментального плана. Эта проблема была решена путем проведения лечения сразу после тренировки с низкой и высокой интенсивностью сотрясения стопы, что позволило избежать индукции зависимости от состояния. Эта манипуляция подтвердила, что инактивация IL препятствует консолидации памяти, когда тренировка IA выполняется с отталкивающим стимулом низкой интенсивности, и показала, что действительно интенсивная тренировка защищает от амнестического эффекта инактивации, производимого TTX (Рисунок 3D).
Заключение
Данные, полученные в этой экспериментальной серии, показывают, что (а) консолидация памяти IA не зависит от нейронной активности ACC; (б) нормальная активность PL важна для консолидации памяти при умеренной тренировке IA, но не для приобретения или закрепления интенсивной тренировки; (c) нормальная активность IL также важна для консолидации памяти при умеренной тренировке IA, но не для приобретения или закрепления интенсивной тренировки. Более того, комбинированный эффект TTX и интенсивных тренировок вызывает зависимость от состояния.
Авторские взносы
Разработал эксперименты: МТ-Г, АМ и РП-А. Проведены поведенческие и гистологические эксперименты: МТ-Г и АМ. Проанализированы данные: МТ-Г и АМ. Написал и предоставил комментарии и обсуждение рукописи: MT-G, AM, GQ и RP-A.
Финансирование
Это исследование было поддержано Генеральным директором Асунтос дель Персональной Академии, Национальным автономным университетом Мексики (UNAM; IN201415) и Consejo Nacional de Ciencia y Tecnología (CONACYT; 237570).MT-G — докторант программы Programa de Doctorado en Ciencias Biomédicas, UNAM и получил стипендию 245656 от CONACYT.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят Берту Ислас, Норму Серафин, Омара Гонсалеса, Леонор Казанову, Мартина Гарсиа и Сандру Эрнандес за их отличную техническую и административную помощь.Мы также хотим поблагодарить Джессику Гонсалес Норрис за редакторские комментарии.
Сноски
- Интенсивная или усиленная тренировка относится к условиям, в которых проводится большое количество попыток или тренировок, и, в случае аверсивного кондиционирования, к тренировкам, мотивированным относительно высокой интенсивностью ударов стопы. В обоих случаях интенсивная тренировка дает более сильное сопротивление исчезновению, чем умеренная и низкая тренировка.
- http: // rsb.info.nih.gov/ij/
Список литературы
Биссьер, С., Плахта, Н., Хойер, Д., Макаллистер, К. Х., Ольпе, Х. Р., Грейс, А. А., и др. (2008). Ростральная часть коры передней поясной извилины модулирует эффективность обучения страху в зависимости от миндалины. Biol. Психиатрия 63, 821–831. DOI: 10.1016 / j.biopsych.2007.10.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Блюм С., Хеберт А. Э. и Дэш П. К. (2006). Роль префронтальной коры в воспроизведении недавних и далеких воспоминаний. Neuroreport 17, 341–344. DOI: 10.1097 / 01.wnr.0000201509.53750.bc
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чай, С. К., Кунг, Дж. К., и Шью, Б. С. (2010). Роль передней поясной коры и медиального таламуса в краткосрочной и долгосрочной обработке аверсивной информации. Mol. Боль 6:42. DOI: 10.1186 / 1744-8069-6-42
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кобос-Сапиаин, Г. Г., Саладо-Кастильо, Р., Санчес-Алавес, М., Кирарте, Г. Л., Рольдан-Рольдан, Г., и Прадо-Алькала, Р. А. (1996). Высокий уровень ударов стопой во время тренировки с ингибирующим избеганием предотвращает амнезию, вызванную интранагральной инъекцией антагонистов ГАМК. Neurobiol. Учить. Mem. 65, 202–206. DOI: 10.1006 / nlme.1996.0024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коркоран, К. А., Квирк, Г. Дж. (2007). Активность предлимбической коры головного мозга необходима для выражения усвоенных, но не врожденных страхов. J. Neurosci. 27, 840–844. DOI: 10.1523 / JNEUROSCI.5327-06.2007
CrossRef Полный текст | Google Scholar
Крус-Моралес, С. Э., Дуран-Аревало, М., Диас дель Гуанте, М. А., Кирарте, Г. Л., и Прадо Алькала, Р. А. (1992). Порог защитного эффекта чрезмерно усиленного пассивного избегания против амнезии, вызванной скополамином. Behav. Neural Biol. 57, 256–259. DOI: 10.1016 / 0163-1047 (92)
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Диас-Трухильо, А., Контрерас, Дж., Медина, А. К., Сильвейра-Леон, Г. А., Кирарте, Г. Л., и Прадо-Алькала, Р. А. (2009). Усиленное обучение избеганию ингибирования предотвращает долговременное ухудшение памяти эффектов циклогексимида, ингибитора синтеза белка. Neurobiol. Учить. Mem. 91, 310–314. DOI: 10.1016 / j.nlm.2008.10.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дуран-Аревало, М., Крус-Моралес, С. Э., и Прадо-Алькала, Р. А. (1990). Участвует ли ацетилхолин в консолидации памяти при чрезмерно усиленном обучении? Brain Res.Бык. 24, 725–727. DOI: 10.1016 / 0361-9230 (90)
-NCrossRef Полный текст | Google Scholar
Галиндо, Л. Е., Гарин-Агилар, М. Е., Медина, А. К., Серафин, Н., Кирарте, Г. Л., и Прадо-Алькала, Р. А. (2008). На приобретение и сохранение усиленного активного избегания не влияет вмешательство в серотонинергическую активность. Behav. Brain Res. 195, 153–158. DOI: 10.1016 / j.bbr.2008.01.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гарин-Агилар, М.Э., Медина, А. С., Кирарте, Г. Л., Макгоу, Дж. Л., и Прадо-Алькала, Р. А. (2014). Интенсивные отталкивающие тренировки защищают память от амнестических эффектов инактивации гиппокампа. Гиппокамп 24, 102–112. DOI: 10.1002 / hipo.22210
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джордано М. и Прадо-Алькала Р. А. (1986). Ретроградная амнезия, вызванная введением атропина в хвостатую скорлупу после исследования. Защитный эффект отрицательного подкрепления. Pharmacol. Biochem. Behav. 24, 905–909. DOI: 10.1016 / 0091-3057 (86)
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джустино Т. Ф. и Марен С. (2015). Роль медиальной префронтальной коры в формировании и подавлении страха. Фронт. Behav. Neurosci. 9: 298. DOI: 10.3389 / fnbeh.2015.00298
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Heidbreder, C.A., и Groenewegen, H.J. (2003).Медиальная префронтальная кора у крысы: доказательства дорсо-вентральной разницы, основанные на функциональных и анатомических характеристиках. Neurosci. Biobehav. Ред. 27, 555–579. DOI: 10.1016 / j.neubiorev.2003.09.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эррера, Д. Г., и Робертсон, Х. А. (1996). Активация c-fos в мозге. Prog. Neurobiol. 50, 83–107. DOI: 10.1016 / S0301-0082 (96) 00021-4
CrossRef Полный текст | Google Scholar
Гувер, W.Б. и Вертес Р. П. (2007). Анатомический анализ афферентных проекций медиальной префронтальной коры у крысы. Brain Struct. Funct. 212, 149–179. DOI: 10.1007 / s00429-007-0150-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Искьердо И. и Макгоу Дж. Л. (2000). Поведенческая фармакология и ее вклад в молекулярные основы консолидации памяти. Behav. Pharmacol. 11, 517–534. DOI: 10.1097 / 00008877-200011000-00001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джинкс, А.Л. и МакГрегор И. С. (1997). Модуляция поведения, связанного с тревогой, после поражения предлимбической или инфралимбической коры у крыс. Brain Res. 772, 181–190. DOI: 10.1016 / S0006-8993 (97) 00810-X
CrossRef Полный текст | Google Scholar
Лазаро-Муньос, Г., Леду, Дж. Э. и Каин, К. К. (2010). Инструментальное избегание Сидмана изначально зависит от латеральной и базальной миндалевидного тела и сдерживается центральными процессами Павлова, опосредованными миндалевидным телом. Biol. Психиатрия 67, 1120–1127.DOI: 10.1016 / j.biopsych.2009.12.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лехнер, Х.А., Сквайр, Л.Р., и Бирн, Дж. Х. (1999). 100 лет консолидации — вспоминая Мюллера и Пильцекера. Learn. Mem. 6, 77–87.
PubMed Аннотация | Google Scholar
Малин, Э. Л., и Макгоу, Дж. Л. (2006). Дифференциальное участие гиппокампа, передней поясной коры и базолатеральной миндалины в памяти для контекста и удара ногой. Proc. Natl. Акад. Sci. США 103, 1959–1963. DOI: 10.1073 / pnas.05108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Макгоу, Дж. Л. (1966). Зависящие от времени процессы в памяти. Science 153, 1351–1358. DOI: 10.1126 / science.153.3742.1351
CrossRef Полный текст | Google Scholar
Мелло и Соуза, Т., Роэслер, Р., Мадруга, М., де-Пари, Ф., Кеведо, Дж., Родригес, К., и др. (1999). Дифференциальные эффекты посттренировочных инфузий мусцимола и AP5 в различных областях поясной коры головного мозга на удерживание для предотвращения ингибирования у крыс. Neurobiol. Учить. Mem. 72, 118–127. DOI: 10.1006 / NLME.1998.3892
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мюллер Г. Э. и Пильцекер А. (1900). Experimentelle beitrage zur lehre vom gedachtniss. Лейпциг: Дж. А. Барт.
Комитет Национального исследовательского совета (США) по обновлению Руководства по уходу и использованию лабораторных животных (2011 г.). Руководство по уходу и использованию лабораторных животных , 8-е изд. Вашингтон, округ Колумбия: Национальная академия прессы.
Google Scholar
Родитель, М. Б., Томаз, К., и Макгоу, Дж. Л. (1992). Повышенная подготовка к выполнению аверсивно мотивированной задачи ослабляет ухудшающие память эффекты посттренировочных поражений миндалины, вызванных N-метил-D-аспартатом. Behav. Neurosci. 106, 789–797. DOI: 10.1037 / 0735-7044.106.5.789
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Родитель, М. Б., Уэст, М., и Макгоу, Дж. Л. (1994). Память крыс с поражением миндалины, индуцированным через 30 дней после тренировки побега, мотивированной ударом ног, отражает степень первоначальной тренировки. Behav. Neurosci. 108, 1080–1087. DOI: 10.1037 / 0735-7044.108.6.1080
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паксинос, Г., Уотсон, К. (2007). Мозг крысы в стереотаксических координатах , 6-е изд. Берлингтон, Массачусетс: Academic Press.
Google Scholar
Перес-Руис К. и Прадо-Алькала Р. А. (1989). Ретроградная амнезия, вызванная инъекцией лидокаина в полосатое тело: защитный эффект отрицательного подкрепления. Brain Res. Бык. 22, 599–603. DOI: 10.1016 / 0361-9230 (89)
-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Prado-Alcalá, R.A., and Cobos-Zapiaín, G.G. (1977). Дефицит обучения, вызванный холинергической блокадой хвостатого ядра, как функция опыта. Brain Res. 138, 190–196. DOI: 10.1016 / 0006-8993 (77)-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Прадо-Алькала, Р. А. и Кобос-Сапиаин, Г.Г. (1979). Вмешательство хлорида калия в деятельность хвостатого ядра. Доказательства «движущейся» инграммы. Brain Res. 172, 577–583. DOI: 10.1016 / 0006-8993 (79)
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Прадо-Алькала Р. А., Кауфман П. и Москона Р. (1980). Инъекции скополамина и KCL в хвостатую скорлупу. Защита от дефицита обучения, вызванная перетренированностью. Pharmacol. Biochem. Behav. 12, 249–253. DOI: 10.1016 / 0091-3057 (80)
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Прадо-Алькала, Р. А., Медина, А. К., Серафин, Н., Кирарте, Г. Л. (2012). Интенсивные эмоциональные переживания и усиленная тренировка предотвращают потерю памяти, вызванную послетренировочной амнестической обработкой полосатого тела, миндалевидного тела, гиппокампа или черной субстанции. Rev. Neurosci. 23, 501–508. DOI: 10.1515 / revneuro-2012-0061
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кирос, К., Мартинес И., Кирарте Г. Л., Моралес Т., Диас-Синтра С. и Прадо-Алькала Р. А. (2003). Усиленное обучение предотвращению торможения предотвращает ухудшающие память эффекты посттренировочной инактивации гиппокампа. Exp. Brain Res. 153, 400–402. DOI: 10.1007 / s00221-003-1704-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Риеккинен, П., Куйтунен, Дж., И Риеккинен, М. (1995). Влияние инфузий скополамина в переднюю и заднюю части поясной извилины на пассивное избегание и навигацию по водному лабиринту. Brain Res. 685, 46–54. DOI: 10.1016 / 0006-8993 (95) 00422-M
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Саккетти Б., Балди Э., Лоренцини К. А. и Бучерелли К. (2003). Роль неокортекса в консолидации обусловливающих страх воспоминаний у крыс. Exp. Brain Res. 152, 323–328. DOI: 10.1007 / s00221-003-1534-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сагар С. М., Шарп Ф. Р. и Курран Т.(1988). Экспрессия белка c-fos в головном мозге: метаболическое картирование на клеточном уровне. Science 240, 1328–1331. DOI: 10.1126 / science.3131879
CrossRef Полный текст | Google Scholar
Саладо-Кастильо, Р., Санчес-Алавес, М., Кирарте, Г. Л., Мартинес Гарсия, М. И., и Прадо-Алькала, Р. А. (2011). Усиленная тренировка защищает память от амнезии, вызванной одновременной инактивацией миндалины и полосатого тела, миндалины и черной субстанции или полосатого тела и черной субстанции. Фронт. Behav. Neurosci. 5:83. DOI: 10.3389 / fnbeh.2011.00083
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сантос-Андерсон, Р. М., и Руттенберг, А. (1976). Стимуляция медиальной или борозды префронтальной коры головного мозга крысы во время обучения пассивному избеганию выборочно влияет на эффективность удержания. Brain Res. 103, 243–259. DOI: 10.1016 / 0006-8993 (76)-6
CrossRef Полный текст | Google Scholar
Солана-Фигероа, Р., Саладо-Кастильо, Р., Кирарте, Г. Л., Галиндо, Л. Е., и Прадо-Алькала, Р. А. (2002). Тренировка усиленного ингибирующего избегания защищает от амнестического действия п-хлорамфетамина. Life Sci. 71, 391–399. DOI: 10.1016 / S0024-3205 (02) 01684-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тан Дж., Ко, С., Дин, Х. К., Цю, С. С., Каледжесан, А. А., и Чжуо, М. (2005). Павловская память о страхе, вызванная активацией передней поясной коры. Mol. Боль 1: 6. DOI: 10.1186 / 1744-8069-1-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тэтчер, Р. У., и Кимбл, Д. П. (1966). Влияние поражений миндалины на сохранение реакции избегания у перетренированных и не перетренированных крыс. Психономический. Sci. 6, 9–10. DOI: 10.3758 / BF03327931
CrossRef Полный текст | Google Scholar
Вертес, Р. П. (2006). Взаимодействие медиальной префронтальной коры, гиппокампа и средней линии таламуса в эмоциональной и когнитивной обработке у крыс. Неврология 142, 1–20. DOI: 10.1016 / j.neuroscience.2006.06.027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Видаль-Гонсалес, И., Видаль-Гонсалес, Б., Раух, С. Л., и Куирк, Г. Дж. (2006). Микростимуляция выявляет противоположные влияния прелимбической и инфралимбической коры на выражение условного страха. Learn. Mem. 13, 728–733. DOI: 10.1101 / пог.м.306106
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уиллоуби, Дж.О., Маккензи, Л., Медведев, А., Хискок, Дж. Дж. (1997). Индукция Fos вслед за системной каиновой кислотой: ранняя экспрессия в гиппокампе и более поздняя широко распространенная экспрессия коррелировали с судорогами. Неврология 77, 379–392. DOI: 10.1016 / S0306-4522 (96) 00462-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Янг, Ф. К., и Лян, К. С. (2014). Взаимодействие дорсального гиппокампа, медиальной префронтальной коры и прилежащего ядра в формировании памяти о страхе: разница в обучении тормозящего избегания и контекстуальной обусловленности страха. Neurobiol. Учить. Mem. 112, 186–194. DOI: 10.1016 / j.nlm.2013.07.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан Ю., Фукусима Х., Кида С. (2011). Индукция и требование экспрессии генов в передней части поясной извилины и медиальной префронтальной коре для консолидации тормозящей памяти избегания. Mol. Мозг 4: 4. DOI: 10.1186 / 1756-6606-4-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Усиленное обучение тормозящего избегания предотвращает ухудшающие память эффекты посттренировочной инактивации гиппокампа
Амброджи Лоренцини К.Г., Балди Э., Бучерелли С., Саккетти Б., Тассони Г. (1999) Нейронная топография и хронология консолидации памяти: обзор функциональных результаты инактивации.Neurobiol Learn Mem 71: 1−18
PubMed Google Scholar
Boehnke S, Rasmusson D (2001) Динамика и эффективное распространение лидокаина и тетродотоксина, доставляемых с помощью микродиализа: электрофизиологическое исследование в коре головного мозга. J Neurosci Methods 105: 133–141
CAS PubMed Google Scholar
Cobos-Zapiaín GG, Salado-Castillo R, Sánchez-Alavez M, Quirarte GL, Roldán-Roldán G, Prado-Alcalá RA (1996) Высокий уровень шока стопы во время тренировки ингибирующего избегания предотвращает амнезию, вызванную интранигральными антагонистами ГАМК .Neurobiol Learn Mem 65: 202–206
Статья PubMed Google Scholar
Джордано М., Прадо-Алькала Р.А. (1986) Ретроградная амнезия, вызванная введением атропина в хвостатую скорлупу после испытания. Защитный эффект отрицательного подкрепления. Pharmacol Biochem Behav 24: 905–909
CAS PubMed Google Scholar
Herrera D, Robertson H (1996) Активация c- fos в головном мозге.Progr Neurobiol 50: 83–107
Статья CAS Google Scholar
Izquierdo I, Da Cunha C, Rosat R, Jerusalinsky D, Ferreira MBC, Medina JH (1992) Рецепторы нейротрансмиттеров, участвующие в обработке памяти миндалевидным телом, медиальной перегородкой и гиппокампом крыс. Behav Neural Biol 58: 16–25
CAS PubMed Google Scholar
Lorenzini CA, Baldi E, Bucherelli C, Sacchetti B, Tassoni G (1996) Роль дорсального гиппокампа в приобретении, консолидации и извлечении реакции пассивного избегания у крыс: исследование функциональной инактивации тетродотоксина.Brain Res 730: 32–39
PubMed Google Scholar
Мартинес I, Quirarte GL, Díaz-Cintra S, Quiroz C, Prado-Alcalá RA (2002) Эффекты поражения полей CA1 и CA3 гиппокампа на приобретение ингибиторного избегания. Нейропсихобиология 46: 97–103
Статья PubMed Google Scholar
Родитель М.Б., Макгоу Дж.Л. (1994) Посттренировочная инфузия лидокаина в базолатеральный комплекс миндалины ухудшает удержание, если тренируется тормозящее избегание.Brain Res 661: 97–103
Статья CAS PubMed Google Scholar
Parent MB, Tomaz C, McGaugh JL (1992) Усиленное обучение выполнению аверсивно мотивированной задачи ослабляет ухудшающие память эффекты посттренировочных поражений миндалины, вызванных N -метил-d-аспартатом. Behav Neurosci 106: 789–797
Статья CAS PubMed Google Scholar
Родитель М.Б., Вест М., Макгоу Дж. Л. (1994) Память крыс с поражением миндалины, индуцированным через 30 дней после тренировки побега, вызванной ударом стопы, отражает степень первоначальной подготовки.Behav Neurosci 108: 1080–1087
Статья CAS PubMed Google Scholar
Paxinos G, Watson C (1982) Мозг крысы в стереотаксических координатах. Academic, New York
Pérez-Ruiz C, Prado-Alcalá R A (1989) Ретроградная амнезия, вызванная инъекцией лидокаина в полосатое тело: защитный эффект отрицательного подкрепления. Brain Res Bull 22: 599–603
CAS PubMed Google Scholar
Prado-Alcalá RA (1995) Последовательная и параллельная обработка во время консолидации памяти.В: McGaugh JL, Bermúdez-Rattoni F, Prado-Alcalá RA (ред.) Пластичность в центральной нервной системе. Обучение и память. Лоуренс Эрлбаум, Нью-Джерси, стр. 57–65
Стубли-Уизерли Л., Хардинг Дж. У., Ррай Дж. У. (1996) Влияние дискретных поражений гиппокампа, вызванных каиновой кислотой, на пространственное и контекстное обучение и память у крыс. Brain Res 716: 29–38
Статья PubMed Google Scholar
Уиллоуби Дж., Маккензи Л., Медведев А., Хискок Дж. (1997) Индукция Fos после системной каиновой кислоты: ранняя экспрессия в гиппокампе и более поздняя широко распространенная экспрессия коррелировали с судорогами.Неврология 77: 379–392
Статья CAS PubMed Google Scholar
Журавин Ю., Бурес Дж. (1991) Степень блокады, вызванной тетродотоксином, исследовали при параличе зрачков, вызванном внутримозговым введением препарата. Exp Brain Res 83: 687–690
CAS PubMed Google Scholar
Система | Объем (л) | Тип |
Смазка (с фильтром) | ||
1GR-FE | 5.2 | API SL..SM |
5VZ-FE | 5,4 | API SL |
2TR-FE | 5,7 | API SL..SM |
1KD-FTV | 7,4 | API CD..CF-4 |
Охлаждение | ||
1GR-FE | 9.8 | |
5VZ-FE | 7,6 | |
2TR-FE | 7,1 | |
1KD-FTV | 10,8 / 11,0 | стандарт / холодная зона |
АКПП | ||
1GR-FE | 10.4 | Тойота ATF WS |
5VZ-FE | 10,1 | Toyota ATF Тип T-IV |
2TR-FE | 10,1 | Toyota ATF Тип T-IV |
1KD-FTV | 10,1 | Toyota ATF Тип T-IV |
Передача | ||
1.4 | Toyota Gear Oil Super GL-5 75W-90 | |
Передний дифференциал | ||
1,4 | Toyota гипоидное трансмиссионное масло SX GL-5 85W-90 | |
Задний дифференциал | ||
std (3d / 5d) | 2,50 / 3,05 | Toyota гипоидное трансмиссионное масло SX GL-5 85W-90 |
Разн.Замок (3д / 5д) | 2,40 / 2,95 | Toyota гипоидное трансмиссионное масло SX GL-5 85W-90 |
LSD (3d / 5d) | 2,50 / 3,05 | Toyota гипоидное трансмиссионное масло LSD GL-5 85W-90 |
Усилитель руля | ||
1GR-FE | 1,0 | Жидкость для гидроусилителя руля Toyota |
5VZ-FE | 0.8 | Жидкость для гидроусилителя руля Toyota |
2TR-FE | 0,7 | Жидкость для гидроусилителя руля Toyota |
1KD-FTV | 1,0 | Жидкость для гидроусилителя руля Toyota |
Бачок омывателя | ||
4.5 | ||
Топливный бак | ||
1GR-FE | 87 | Премиум (95 RON) |
5VZ-FE, 2TR-FE | 87 | Обычный (RON 91) |
1KD-FTV | 87 | Дизель |
Модель мыши при врожденной миастении
% PDF-1.7 % 1 0 obj > / OCGs [6 0 R] >> / Контуры 7 0 R / Страницы 3 0 R / Тип / Каталог >> эндобдж 2 0 obj > поток 2019-07-29T06: 32: 27-07: 002019-07-29T06: 32: 27-07: 002019-07-29T06: 32: 27-07: 00Appligent AppendPDF Pro 5.5uuid: 44bc37f2-ac1a-11b2-0a00- 782dad000000uuid: 44bc69e9-ac1a-11b2-0a00-f0e69951fd7fapplication / pdf
Молекулярные механизмы, лежащие в основе действия простагландинов, производных арахидоновой кислоты, на периферическую ноцицепцию | Журнал нейровоспаления
Деннис EA, Норрис PC. Эйкозаноидный шторм при инфекции и воспалении. Nat Rev Immunol. 2015; 15: 511–23.
CAS PubMed PubMed Central Статья Google Scholar
Пето Г., Рих П.В.Сенсорные и сигнальные механизмы брадикинина, эйкозаноидов, фактора активации тромбоцитов и оксида азота в периферических ноцицепторах. Physiol Rev.2012; 92: 1699–775.
CAS PubMed Статья Google Scholar
Straus DS, Glass CK. Циклопентеноновые простагландины: новое понимание биологической активности и клеточных мишеней. Med Res Rev.2001; 21: 185–210.
CAS PubMed Статья Google Scholar
Брейер Р.М., Багдассарян СК, Майерс С.А., Брейер М.Д. Простаноидные рецепторы: подтипы и сигнализация. Annu Rev Pharmacol Toxicol. 2001; 41: 661–90.
CAS PubMed Статья Google Scholar
Кавамура М., Инаока Х., Обата С., Харада Ю. Почему многие животные сохраняют несколько изоформ циклооксигеназы? Простагландины Other Lipid Mediat. 2014; 109–111: 14–22.
CAS PubMed Статья Google Scholar
Самад Т.А., Мур К.А., Сапирштейн А., Биллет С., Олчорн А., Пул С., Бонвентре СП, Вульф С.Дж. Опосредованная интерлейкином-1бета индукция ЦОГ-2 в ЦНС способствует воспалительной гиперчувствительности к боли. Природа. 2001; 410: 471–5.
CAS PubMed Статья Google Scholar
Beiche F, Scheuerer S, Brune K, Geisslinger G, Goppelt-Struebe M. Повышение регуляции мРНК циклооксигеназы-2 в спинном мозге крысы после периферического воспаления.FEBS Lett. 1996; 390: 165–9.
CAS PubMed Статья Google Scholar
Ebersberger A, Grubb BD, Willingale HL, Gardiner NJ, Nebe J, Schaible HG. Интраспинальное высвобождение простагландина E2 в модели острого артрита сопровождается повышающей регуляцией циклооксигеназы-2 в спинном мозге. Неврология. 1999; 93: 775–81.
CAS PubMed Статья Google Scholar
Тегедер I, Нидербергер Э., Веттер Г., Браутигам Л., Гейсслингер Г. Влияние избирательного ингибирования ЦОГ-1 и -2 на вызываемое формалином ноцицептивное поведение и выброс простагландина E (2) в спинном мозге. J Neurochem. 2001; 79: 777–86.
CAS PubMed Статья Google Scholar
Якш Т.Л., Дириг Д.М., Конвей К.М., Свенссон К., Луо З.Д., Исаксон П.С. Острое антигипералгезическое действие нестероидных противовоспалительных препаратов и высвобождение простагландина E2 спинного мозга опосредовано ингибированием конститутивной спинномозговой циклооксигеназы-2 (ЦОГ-2), но не ЦОГ-1.J Neurosci. 2001; 21: 5847–53.
CAS PubMed PubMed Central Статья Google Scholar
Хефферан М.П., Картер П., Хейли М., Лумис К.В. Повреждение спинного нерва активирует синтез простагландинов в спинном мозге, что способствует раннему поддержанию тактильной аллодинии. Боль. 2003. 101: 139–47.
CAS PubMed Статья Google Scholar
Zhu X, Eisenach JC. Циклооксигеназа-1 в спинном мозге изменяется после повреждения периферического нерва. Анестезиология. 2003; 99: 1175–9.
CAS PubMed Статья Google Scholar
Гриль М., Пескар Б.А., Шулигой Р., Аманн Р. Системное воспаление индуцирует опосредованный ЦОГ-2 биосинтез простагландина D2 в спинном мозге мышей. Нейрофармакология. 2006; 50: 165–73.
CAS PubMed Статья Google Scholar
Виллингейл HL, Гардинер Нью-Джерси, Маклаймонт Н, Гиблетт С., Грабб Б.Д. Простаноиды, синтезируемые изоформами циклооксигеназы в спинном мозге крыс, и их вклад в развитие гипервозбудимости нейронов. Br J Pharmacol. 1997; 122: 1593–604.
CAS PubMed PubMed Central Статья Google Scholar
Баллоу Л.Р., Боттинг Р.М., Гурха С., Чжан Дж., Вейн Дж. Ноцицепция у мышей с дефицитом изофермента циклооксигеназы.Proc Natl Acad Sci U S. A. 2000; 97: 10272–6.
CAS PubMed PubMed Central Статья Google Scholar
Чопра Б., Гиблетт С., Литтл Дж. Г., Дональдсон Л. Ф., Тейт С., Эванс Р. Дж., Грабб Б. Д.. Циклооксигеназа-1 является маркером субпопуляции предполагаемых ноцицептивных нейронов в ганглиях задних корешков крыс. Eur J Neurosci. 2000; 12: 911–20.
CAS PubMed Статья Google Scholar
Dou W, Jiao Y, Goorha S, Raghow R, Ballou LR. Ноцицепция и дифференциальная экспрессия циклооксигеназы-1 (COX-1), варианта COX-1, сохраняющего интрон-1 (COX-1v), и COX-2 в ганглиях задних корешков мыши (DRG). Простагландины Other Lipid Mediat. 2004. 74: 29–43.
CAS PubMed Статья Google Scholar
Гриль М., Хайнеманн А., Хефлер Г., Пескар Б.А., Шулигой Р. Влияние лечения эндотоксинами на экспрессию и локализацию спинномозговой циклооксигеназы, простагландинсинтаз и рецепторов PGD2.J Neurochem. 2008. 104: 1345–57.
CAS PubMed Статья Google Scholar
Фам-Марку Т.А., Белойл Х., Сан Х, Джентили М., Яичи Д., Бенуа Дж., Бенхаму Д., Мазойт Дж. Х. Антиноцицептивный эффект ресвератрола при гипералгезии, вызванной каррагенаном, у крыс: пролонгированный эффект, связанный с нарушением экспрессии ЦОГ-2. Боль. 2008. 140: 274–83.
CAS PubMed Статья Google Scholar
Амайя Ф, Самад Т.А., Барретт Л., Брум, округ Колумбия, Вульф СиДжей. Периганглионарное воспаление вызывает гиперчувствительность к боли, излучающую дистально, способствуя индукции ЦОГ-2 в ганглии дорсального корешка. Боль. 2009. 142: 59–67.
CAS PubMed PubMed Central Статья Google Scholar
Araldi D, Ferrari LF, Lotufo CM, Vieira AS, Athie MC, Figueiredo JG, Duarte DB, Tambeli CH, Ferreira SH, Parada CA. Периферическая воспалительная гипералгезия зависит от увеличения ЦОГ в ганглии задних корешков.Proc Natl Acad Sci U S. A. 2013; 110: 3603–8.
CAS PubMed PubMed Central Статья Google Scholar
Le Pichon CE, Chesler AT. Функциональное и анатомическое рассечение соматосенсорных субпопуляций с использованием генетики мышей. Фронт нейроанат. 2014; 8:21.
PubMed PubMed Central Статья CAS Google Scholar
Чан Й, Чо ПС, Ян Й. Д., Хван, SW.Ноцицептивная роль ионного канала TRPM2 при патологической боли. Mol Neurobiol. 2018; 55: 6589–600.
CAS PubMed Статья Google Scholar
Inoue A, Iwasa M, Nishikura Y, Ogawa S, Nakasuka A, Nakata Y. Длительное воздействие брадикинина на культивированные на крысах клетки ганглиев дорзального корешка индуцировало высвобождение простагландина E2 за счет активации циклооксигеназы. 2. Neurosci Lett. 2006; 401: 242–7.
CAS PubMed Статья Google Scholar
Cheng L, Jia H, Lohr M, Bagherzadeh A, Holmes D.I, Selwood D, Zachary I. Антихеморепульсивные эффекты фактора роста эндотелия сосудов и фактора роста плаценты-2 в нейронах ганглиев дорзального корешка опосредуются нейропилином-1 и циклооксигеназой. -производная простаноидная продукция. J Biol Chem. 2004. 279: 30654–61.
CAS PubMed Статья Google Scholar
Веллани В., Франки С., Прандини М., Моретти С., Кастелли М., Джакомони К., Сакердот П.Влияние НПВП и парацетамола (ацетаминофена) на транслокацию протеинкиназы C-эпсилон, а также на синтез и высвобождение вещества P в культивируемых сенсорных нейронах. J Pain Res. 2013; 6: 111–20.
CAS PubMed PubMed Central Статья Google Scholar
Ma K, Zhou QH, Chen J, Du DP, Ji Y, Jiang W. Снижение тока TTX-R Na + под действием целекоксиба коррелирует с изменениями в PGE (2) и CGRP в нейронах DRG крыс во время острого послеоперационного вмешательства. боль.Brain Res. 2008; 1209: 57–64.
CAS PubMed Статья Google Scholar
Ван И, Чжан Х, Гуо QL, Цзоу Вайоминг, Хуанг К.С., Ян Дж. Ингибиторы циклооксигеназы подавляют экспрессию рецепторов P2X (3) в DRG и ослабляют гипералгезию после хронического сужения у крыс. Neurosci Lett. 2010; 478: 77–81.
CAS PubMed Статья Google Scholar
Syriatowicz JP, Hu D, Walker JS, Tracey DJ. Гипералгезия из-за повреждения нерва: роль простагландинов. Неврология. 1999; 94: 587–94.
CAS PubMed Статья Google Scholar
Goncalves MB, Williams EJ, Yip P, Yanez-Munoz RJ, Williams G, Doherty P. Ингибиторы ЦОГ-2, мелоксикам и нимесулид подавляют нейрогенез в мозге взрослых мышей. Br J Pharmacol. 2010; 159: 1118–25.
CAS PubMed PubMed Central Статья Google Scholar
Fehrenbacher JC, Burkey TH, Nicol GD, Vasko MR. Фактор некроза опухоли альфа и интерлейкин-1бета стимулируют экспрессию циклооксигеназы II, но не изменяют уровни мРНК рецептора простагландина E2 в культивируемых клетках ганглиев задних корешков. Боль. 2005; 113: 113–22.
CAS PubMed Статья Google Scholar
Gould HJ 3rd, England JD, Soignier RD, Nolan P, Minor LD, Liu ZP, Levinson SR, Paul D. Ибупрофен блокирует изменения Na v 1.7 и 1.8 натриевые каналы связаны с полным воспалением, вызванным адъювантом Фрейнда, у крыс. J Pain. 2004; 5: 270–80.
CAS PubMed Статья Google Scholar
Иноуэ А, Икома К., Мориока Н., Кумагаи К., Хашимото Т., Хиде I, Наката Ю. Интерлейкин-1бета индуцирует высвобождение вещества Р из первичных афферентных нейронов через систему циклооксигеназы-2. J Neurochem. 1999; 73: 2206–13.
CAS PubMed Google Scholar
Morioka N, Inoue A, Hanada T, Kumagai K, Takeda K, Ikoma K, Hide I, Tamura Y, Shiomi H, Dohi T, Nakata Y. Оксид азота синергетически усиливает индуцированное интерлейкином-1 бета увеличение мРНК циклооксигеназы-2 уровни, приводящие к облегчению высвобождения вещества P из первичных афферентных нейронов: участие cGMP-независимых механизмов. Нейрофармакология. 2002; 43: 868–76.
CAS PubMed Статья Google Scholar
Cruz Duarte P, St-Jacques B, Ma W. Простагландин E2 способствует синтезу нейротрофического фактора головного мозга в первичных сенсорных нейронах в культурах эксплантатов ганглиев и в модели нейропатической боли. Exp Neurol. 2012; 234: 466–81.
CAS PubMed Статья Google Scholar
Юнгер Х., Соркин Л.С. Сенсибилизация C-ноцицептора изопростанами зависит от циклооксигеназы. Brain Res. 2000; 867: 255–8.
CAS PubMed Статья Google Scholar
Николь Г. Д., Лопшир Дж. С., Паффорд К. М.. Фактор некроза опухоли увеличивает чувствительность сенсорных нейронов крысы к капсаицину. J Neurosci. 1997; 17: 975–82.
CAS PubMed PubMed Central Статья Google Scholar
Джанг И.Дж., Дэвис А.Дж., Акимото Н., Бэк С.К., Ли ПР, На ХС, Фуру Х., Юнг С.Дж., Ким Й.Х., О С.Б. Острое воспаление выявляет опосредованную рецептором GABAA ноцицепцию в нейронах ганглия задних корешков мышей посредством передачи сигналов рецептора 4 PGE2.Physiol Rep.2017; 5: e13178.
PubMed PubMed Central Статья CAS Google Scholar
Sun W, Yang F, Wang Y, Fu H, Yang Y, Li CL, Wang XL, Lin Q, Chen J. Вклад первичной сенсибилизации сенсорных нейронов большого размера в механическую аллодинию за счет активации гиперполяризации. активирует закрытые циклическими нуклеотидами каналы через каскад циклооксигеназы 1. Нейрофармакология. 2017; 113: 217–30.
CAS PubMed Статья Google Scholar
Taiwo Y, Heller P, Levine J. Характеристика различных фосфолипаз, опосредующих гипералгезию брадикинина и норадреналина. Неврология. 1990; 39: 523–31.
CAS PubMed Статья Google Scholar
Феррейра С., Лоренцетти Б., Пул С. Брадикинин инициирует опосредованную цитокинами воспалительную гипералгезию. Br J Pharmacol. 1993; 110: 1227–31.
CAS PubMed PubMed Central Статья Google Scholar
Supowit SC, Zhao H, Katki KA, Gupta P, Dipette DJ. Брадикинин и простагландин E (1) регулируют экспрессию пептидов, связанных с геном кальцитонина, в культивируемых сенсорных нейронах крыс. Regul Pept. 2011; 167: 105–11.
CAS PubMed Статья Google Scholar
Дженкинс Д.В., Селлерс Л.А., Фенюк В., Хамфри П.П. Характеристика индуцированного брадикинином высвобождения простагландина E2 из культивированных нейронов тройничного ганглия крыс. Eur J Pharmacol.2003. 469: 29–36.
CAS PubMed Статья Google Scholar
Xanthos DN, Sandkuhler J. Нейрогенное нейровоспаление: воспалительные реакции ЦНС в ответ на активность нейронов. Nat Rev Neurosci. 2014; 15: 43–53.
CAS PubMed Статья Google Scholar
Ma W, Chabot JG, Vercauteren F, Quirion R. Поврежденный СОХ2 / PGE2 нервного происхождения способствует поддержанию невропатической боли у старых крыс.Neurobiol Aging. 2010; 31: 1227–37.
CAS PubMed Статья Google Scholar
Murakami M, Naraba H, Tanioka T, Semmyo N, Nakatani Y, Kojima F, Ikeda T, Fueki M, Ueno A, Oh S, Kudo I. Регулирование биосинтеза простагландина E2 индуцируемым мембранно-ассоциированным простагландином Е2-синтаза, действующая совместно с циклооксигеназой-2. J Biol Chem. 2000; 275: 32783–92.
CAS PubMed Статья Google Scholar
Urade Y, Eguchi N. Липокалин-тип и гемопоэтические простагландин D-синтазы как новый пример функциональной конвергенции. Простагландины Other Lipid Mediat. 2002; 68-69: 375-82.
CAS PubMed Статья Google Scholar
Ujihara M, Urade Y, Eguchi N, Hayashi H, Ikai K, Hayaishi O. Образование простагландина D2 и характеристика его синтетаз в различных тканях взрослых крыс. Arch Biochem Biophys. 1988; 260: 521–31.
CAS PubMed Статья Google Scholar
Pinho-Ribeiro FA, Verri WA Jr, Chiu IM. Ноцицепторные сенсорные нейронно-иммунные взаимодействия при боли и воспалении. Trends Immunol. 2017; 38: 5–19.
CAS PubMed Статья Google Scholar
Cunha TM, Verri WA Jr, Schivo IR, Napimoga MH, Parada CA, Poole S, Teixeira MM, Ferreira SH, Cunha FQ.Решающая роль нейтрофилов в развитии механической воспалительной гиперноцицепции. J Leukoc Biol. 2008; 83: 824–32.
CAS PubMed Статья Google Scholar
Ма В., Квирион Р. Повышающая регуляция интерлейкина-6, индуцированная простагландином Е, из-за вторжения макрофагов после повреждения нерва: исследование in vivo и in vitro. J Neurochem. 2005; 93: 664–73.
CAS PubMed Статья Google Scholar
Чопра С., Джованелли П., Альварадо-Васкес П.А., Алонсо С., Сонг М., Сандовал Т.А., Чае С.С., Тан С., Фонсека М.М., Гутьеррес С. и др. Передача сигналов IRE1alpha-XBP1 в лейкоцитах контролирует биосинтез простагландинов и боль. Наука. 2019; 365: eaau6499.
CAS PubMed Статья Google Scholar
Весин М.Ф., Дроз Б. Биосинтез простагландинов D2 и E2 в ганглиях дорсального корешка цыпленка во время развития. J Neurochem. 1991; 57: 161–6.
CAS PubMed Статья Google Scholar
Весин М.Ф., Баракат-Вальтер И., Дроз Б. Предпочтительный синтез простагландина D2 нейронами и простагландина E2 фибробластами и ненейрональными клетками в ганглиях задних корешков кур. J Neurochem. 1991; 57: 167–74.
CAS PubMed Статья Google Scholar
Весин М.Ф., Ураде Y, Хаяиши О, Дроз Б.Нейрональные и глиальные изоферменты простагландин D-синтазы в ганглиях задних корешков кур: иммуноцитохимическое исследование с помощью светового и электронного микроскопа. J Neurosci. 1995; 15: 470–6.
CAS PubMed PubMed Central Статья Google Scholar
Весин М.Ф., Дроз Б. Иммунодетекция простагландин-D-синтазы: условия локализации в определенном подклассе первичных сенсорных нейронов. J Histochem Cytochem. 1995; 43: 681–7.
CAS PubMed Статья Google Scholar
Schuligoi R, Ulcar R, Peskar BA, Amann R. Влияние лечения эндотоксином на экспрессию циклооксигеназы-2 и простагландин-синтаз в спинном мозге, ганглиях задних корешков и коже крыс. Неврология. 2003; 116: 1043–52.
CAS PubMed Статья Google Scholar
Кунори С., Мацумура С., Окуда-Ашитака Е., Катано Т., Аудоли Л. П., Ураде Ю., Ито С. Новая роль простагландина Е2 в невропатической боли: блокада миграции микроглии в спинном мозге.Глия. 2011; 59: 208–18.
PubMed Статья Google Scholar
Trimarco A, Forese MG, Alfieri V, Lucente A, Brambilla P, Dina G, Pieragostino D, Sacchetta P, Urade Y, Boizet-Bonhoure B и др. Простагландин D2-синтаза / GPR44: сигнальная ось миелинизации ПНС. Nat Neurosci. 2014; 17: 1682–92.
CAS PubMed Статья Google Scholar
Шаферс М., Марзиняк М., Соркин Л.С., Якш Т.Л., Соммер К.Ингибирование циклооксигеназы при гипералгезии, вызванной повреждением нервов и TNF, у крыс. Exp Neurol. 2004. 185: 160–8.
PubMed Статья CAS Google Scholar
Souza GR, Talbot J, Lotufo CM, Cunha FQ, Cunha TM, Ferreira SH. Фракталкин опосредует воспалительную боль за счет активации сателлитных глиальных клеток. Proc Natl Acad Sci U S. A. 2013; 110: 11193–8.
CAS PubMed PubMed Central Статья Google Scholar
Zeisel A, Hochgerner H, Lonnerberg P, Johnsson A, Memic F, van der Zwan J, Haring M, Braun E, Borm LE, La Manno G, et al. Молекулярная архитектура нервной системы мыши. Клетка. 2018; 174: 999–1014 e1022.
CAS PubMed PubMed Central Статья Google Scholar
Остин П.Дж., Моалем-Тейлор Г. Нейроиммунный баланс при нейропатической боли: участие воспалительных иммунных клеток, иммуноподобных глиальных клеток и цитокинов.J Neuroimmunol. 2010; 229: 26–50.
CAS PubMed Статья Google Scholar
Гиларди Дж. Р., Свенссон К. И., Роджерс С. Д., Якш Т. Л., Манты П. В.. Конститутивная спинномозговая циклооксигеназа-2 участвует в инициации гипералгезии, вызванной повреждением тканей. J Neurosci. 2004. 24: 2727–32.
CAS PubMed PubMed Central Статья Google Scholar
Свенссон К.И., Хуа XY, Проттер А.А., Пауэлл Х.С., Якш, TL. Спинальная киназа p38 MAP необходима для индуцированного NMDA высвобождения спинномозгового PGE (2) и тепловой гипералгезии. Нейроотчет. 2003. 14: 1153–7.
CAS PubMed Статья Google Scholar
Дириг Д.М., Якшская обл. Спинальный синтез и высвобождение простаноидов после периферических повреждений и воспалений. Adv Exp Med Biol. 1999; 469: 401–8.
CAS PubMed Статья Google Scholar
Muja N, DeVries GH. Продукция простагландина E (2) и 6-кето-простагландина F (1альфа) повышается после травматического повреждения седалищного нерва. Глия. 2004; 46: 116–29.
PubMed Статья Google Scholar
Capuano A, De Corato A, Lisi L, Tringali G, Navarra P, Dello Russo C. Активированные воспалением сателлитные клетки тройничного нерва способствуют сенсибилизации нейронов: актуальность для патологии мигрени. Молочная боль. 2009; 5: 43.
PubMed PubMed Central Статья CAS Google Scholar
Trebino CE, Stock JL, Gibbons CP, Naiman BM, Wachtmann TS, Umland JP, Pandher K, Lapointe JM, Saha S, Roach ML, et al. Нарушение воспалительной и болевой реакции у мышей, лишенных индуцибельной простагландин-E-синтазы. Proc Natl Acad Sci U S. A. 2003; 100: 9044–9.
CAS PubMed PubMed Central Статья Google Scholar
Камей Д., Ямакава К., Такегоши Ю., Миками-Наканиши М., Накатани Ю., О-Иши С., Ясуи Х., Адзума Ю., Хирасава Н., Охучи К. и др. Снижение болевой гиперчувствительности и воспаления у мышей, лишенных микросомальной простагландин-е-синтазы-1. J Biol Chem. 2004. 279: 33684–95.
CAS PubMed Статья Google Scholar
Brenneis C, Coste O, Schmidt R, Angioni C, Popp L, Nusing RM, Becker W., Scholich K, Geisslinger G. Последствия измененных паттернов эйкозаноидов для ноцицептивного процессинга у мышей с дефицитом mPGES-1.J Cell Mol Med. 2008; 12: 639–48.
CAS PubMed Статья Google Scholar
Мабучи Т., Кодзима Х., Абе Т., Такаги К., Сакураи М., Омия Й., Уэмацу С., Акира С., Ватанабе К., Ито С. Простагландин Е-синтаза-1, ассоциированная с мембраной, необходима при невропатической боли. Нейроотчет. 2004; 15: 1395–8.
CAS PubMed Статья Google Scholar
Scholich K, Geisslinger G.Является ли mPGES-1 многообещающей мишенью для лечения боли? Trends Pharmacol Sci. 2006. 27: 399–401.
CAS PubMed Статья Google Scholar
Eguchi N, Minami T, Shirafuji N, Kanaoka Y, Tanaka T, Nagata A, Yoshida N, Urade Y, Ito S, Hayaishi O. Отсутствие тактильной боли (аллодинии) в простагландин D-синтазе липокалинового типа -дефицитные мыши. Proc Natl Acad Sci U S. A. 1999; 96: 726–30.
CAS PubMed PubMed Central Статья Google Scholar
Кавахара К., Ходжох Х., Иназуми Т., Цучия С., Сугимото Ю. Воспаление, вызванное простагландином E2: актуальность рецепторов простагландина E. Biochim Biophys Acta. 1851; 2015: 414–21.
Google Scholar
Jabbour HN, Sales KJ. Передача сигналов и функция рецептора простагландина при патологии эндометрия человека. Trends Endocrinol Metab. 2004; 15: 398–404.
CAS PubMed Статья Google Scholar
Oida H, Namba T, Sugimoto Y, Ushikubi F, Ohishi H, Ichikawa A, Narumiya S. Исследования in situ гибридизации экспрессии мРНК рецептора простациклина в различных органах мыши. Br J Pharmacol. 1995; 116: 2828–37.
CAS PubMed PubMed Central Статья Google Scholar
Саутхолл MD, Васько MR. Подтипы рецепторов простагландина, EP3C и EP4, опосредуют индуцированное простагландином E2 производство цАМФ и сенсибилизацию сенсорных нейронов.J Biol Chem. 2001; 276: 16083–91.
CAS PubMed Статья Google Scholar
Natura G, Bar KJ, Eitner A, Boettger MK, Richter F, Hensellek S, Ebersberger A, Leuchtweis J, Maruyama T., Hofmann GO и др. Подтип EP3 нейронального рецептора простагландина E2 опосредует антиноцицепцию во время воспаления. Proc Natl Acad Sci U S. A. 2013; 110: 13648–53.
CAS PubMed PubMed Central Статья Google Scholar
Patwardhan AM, Vela J, Farugia J, Vela K, Hargreaves KM. Ноцицепторы тройничного нерва экспрессируют рецепторы простагландина. J Dent Res. 2008. 87: 262–6.
CAS PubMed Статья Google Scholar
Крас СП, Донг Л., Винкельштейн Б.А. Рецептор простагландина E2, EP2, активируется в ганглии дорсального корешка после болезненного повреждения шейно-фасеточного сустава у крысы. Позвоночник (Phila Pa 1976). 2013; 38: 217–22.
Артикул Google Scholar
Durrenberger PF, Facer P, Casula MA, Yiangou Y, Gray RA, Chessell IP, Day NC, Collins SD, Bingham S, Wilson AW, et al. Простаноидный рецептор EP1 и cox-2 в поврежденных нервах человека и крысиная модель повреждения нерва: исследование с течением времени. BMC Neurol. 2006; 6: 1.
PubMed PubMed Central Статья CAS Google Scholar
Гривз Э., Хорн А.В., Джерина Х., Миколайчак М., Хилферти Л., Митчелл Р., Флитвуд-Уокер С.М., Сондерс П.Т.Антагонизм рецептора EP2 снижает периферическую и центральную гипералгезию на доклинической модели эндометриоза у мышей. Научный отчет 2017; 7: 44169.
PubMed PubMed Central Статья Google Scholar
Секигучи М., Отоши К., Кикучи С., Конно С. Анальгетические эффекты антагониста рецептора EP1 подтипа рецептора простагландина E2: экспериментальное исследование применения пульпозного ядра. Позвоночник (Phila Pa 1976). 2011; 36: 1829–34.
Артикул Google Scholar
Боргланд С.Л., Коннор М, Райан Р.М., Болл Х.Дж., Кристи МДж. Простагландин E (2) ингибирует ток кальция в двух субпопуляциях остро изолированных сенсорных нейронов тройничного нерва мыши. J Physiol. 2002; 539: 433–44.
CAS PubMed PubMed Central Статья Google Scholar
Бар KJ, Natura G, Telleria-Diaz A, Teschner P, Vogel R, Vasquez E, Schaible HG, Ebersberger A. Изменения действия простагландина E2 спинного мозга при воспалении: простагландин E (EP1-EP4) рецепторы в спинномозговой ноцицептивной обработке сигналов, поступающих от нормального или воспаленного коленного сустава.J Neurosci. 2004. 24: 642–51.
PubMed PubMed Central Статья CAS Google Scholar
Treutlein EM, Kern K, Weigert A, Tarighi N, Schuh CD, Nusing RM, Schreiber Y, Ferreiros N, Brune B., Geisslinger G, et al. Рецептор простагландина E2 EP3 контролирует опосредованную CC-хемокиновым лигандом 2 невропатическую боль, вызванную механическим повреждением нерва. J Biol Chem. 2018; 293: 9685–95.
CAS PubMed PubMed Central Статья Google Scholar
Nakayama Y, Omote K, Kawamata T., Namiki A. Роль подтипа EP1 рецептора простагландина в индуцированной простагландином E2 ноцицептивной передаче в спинном заднем роге крысы. Brain Res. 2004; 1010: 62–8.
CAS PubMed Статья Google Scholar
Gold MS, Levine JD, Correa AM. Модуляция TTX-R INa с помощью PKC и PKA и их роль в PGE2-индуцированной сенсибилизации сенсорных нейронов крысы in vitro. J Neurosci. 1998; 18: 10345–55.
CAS PubMed PubMed Central Статья Google Scholar
Sachs D, Villarreal C, Cunha F, Parada C, Ferreira S. Роль путей PKA и PKCepsilon в опосредованной простагландином E2 гиперноцицепции. Br J Pharmacol. 2009; 156: 826–34.
CAS PubMed PubMed Central Статья Google Scholar
Waters MJ, Brooks AJ. Активация JAK2 гормоном роста и другими цитокинами.Биохим Дж. 2015; 466: 1–11.
CAS PubMed PubMed Central Статья Google Scholar
Тамура С., Морикава Ю., Сенба Е. Активное фосфорилирование сигнального преобразователя и активатора транскрипции 3 и белка, связывающего циклический АМФ-чувствительный элемент, посредством периферического воспаления в первичных афферентных нейронах, возможно, через рецептор онкостатина М. Неврология. 2005; 133: 797–806.
CAS PubMed Статья Google Scholar
Домингес Э., Риват С., Поммье Б., Моборн А., Поль М. Путь JAK / STAT3 активируется в микроглии спинного мозга после повреждения периферического нерва и способствует развитию нейропатической боли у крыс. J Neurochem. 2008; 107: 50–60.
CAS PubMed Статья Google Scholar
Vieira AS, Araldi D, Dias EV, do Prado FC, Tambeli CH, Parada CA. Активация янус-киназы 2 участвует в гипералгезии, вызванной простагландином E2.Life Sci. 2016; 166: 8–12.
CAS PubMed Статья Google Scholar
Алей К.О., Маккартер Дж., Левин Дж. Д.. Передача сигналов оксида азота при боли и сенсибилизации ноцицепторов у крыс. J Neurosci. 1998. 18: 7008–14.
CAS PubMed PubMed Central Статья Google Scholar
St-Jacques B, Ma W. Роль простагландина E2 в синтезе провоспалительного цитокина интерлейкина-6 в первичных сенсорных нейронах: исследование in vivo и in vitro.J Neurochem. 2011; 118: 841–54.
CAS PubMed Статья Google Scholar
Tse KH, Chow KB, Leung WK, Wong YH, Wise H. Липополисахарид по-разному модулирует экспрессию цитокинов и циклооксигеназ в ганглиозных клетках дорсального корешка через toll-подобные рецептор-4 зависимые пути. Неврология. 2014; 267: 241–51.
CAS PubMed Статья Google Scholar
Tse KH, Chow KB, Leung WK, Wong YH, Wise H. Первичные сенсорные нейроны регулируют toll-подобную рецептор-4-зависимую активность глиальных клеток в ганглиях задних корешков. Неврология. 2014; 279: 10–22.
CAS PubMed Статья Google Scholar
Tse KH, Chow KB, Wise H. PGE2, высвобождаемый первичными сенсорными нейронами, модулирует активность toll-подобного рецептора 4 посредством процесса, зависимого от рецептора EP4. J Neuroimmunol. 2016; 293: 8–16.
CAS PubMed Статья Google Scholar
Баба Х, Коно Т., Мур К.А., Вульф С.Дж. Прямая активация нейронов спинного мозга крысы простагландином E2. J Neurosci. 2001; 21: 1750–6.
CAS PubMed PubMed Central Статья Google Scholar
Ahmadi S, Lippross S, Neuhuber WL, Zeilhofer HU. PGE (2) избирательно блокирует ингибирующую глицинергическую нейротрансмиссию на нейроны поверхностного дорсального рога крысы.Nat Neurosci. 2002; 5: 34–40.
CAS PubMed Статья Google Scholar
Харви Р.Дж., Депнер У.Б., Вассл Х., Ахмади С., Хайндл С., Рейнольд Х., Смарт Т.Г., Харви К., Шутц Б., Або-Салем О.М. и др. GlyR alpha3: важная мишень для сенсибилизации боли, опосредованной PGE2. Наука. 2004; 304: 884–7.
CAS PubMed Статья Google Scholar
Reinold H, Ahmadi S, Depner UB, Layh B, Heindl C, Hamza M, Pahl A, Brune K, Narumiya S, Muller U, Zeilhofer HU. Воспалительная гипералгезия позвоночника опосредуется рецепторами простагландина E подтипа EP2. J Clin Invest. 2005; 115: 673–9.
CAS PubMed PubMed Central Статья Google Scholar
Хосл К., Рейнольд Х., Харви Р.Дж., Мюллер У., Нарумия С., Цайльхофер Х. Спинальные рецепторы простагландина E подтипа EP2 и субъединица альфа3 глицинового рецептора, которые опосредуют центральную воспалительную гипералгезию, не вызывают боли после повреждения периферического нерва или инъекции формалина.Боль. 2006; 126: 46–53.
PubMed Статья CAS Google Scholar
Li J, Serafin E, Baccei ML. Простагландиновая передача сигналов управляет зависимой от времени пластичностью спайков в сенсорных синапсах на проекционные нейроны позвоночника мыши. J Neurosci. 2018; 38: 6628–39.
CAS PubMed PubMed Central Статья Google Scholar
Смит Дж. А., Амагасу С. М., Эглен Р. М., Хантер Дж. К., Блей К. Р..Характеристика ответов, вызванных простаноидными рецепторами в сенсорных нейронах крыс. Br J Pharmacol. 1998; 124: 513–23.
CAS PubMed PubMed Central Статья Google Scholar
Роулендс Д.К., Као С., Уайз Х. Регулирование опосредованных рецептором простациклина и простагландина E (2) ответов в ганглиозных клетках дорсального корешка взрослых крыс in vitro. Br J Pharmacol. 2001; 133: 13–22.
CAS PubMed PubMed Central Статья Google Scholar
Wise H. Отсутствие взаимодействия между подтипами рецепторов простагландина E2 в регуляции активности аденилатциклазы в культивируемых клетках ганглиев дорсального корешка крысы. Eur J Pharmacol. 2006; 535: 69–77.
CAS PubMed Статья Google Scholar
Лин Ч.Р., Амая Ф., Барретт Л., Ван Х., Такада Дж., Самад Т.А., Вульф СиДжей. Рецептор простагландина E2 EP4 способствует воспалительной гиперчувствительности к боли. J Pharmacol Exp Ther. 2006; 319: 1096–103.
CAS PubMed Статья Google Scholar
St-Jacques B, Ma W. Передача сигналов простагландина E2 / EP4 способствует экстернализации рецептора EP4 в первичных сенсорных нейронах in vitro и in vivo. Боль. 2013; 154: 313–23.
CAS PubMed Статья Google Scholar
St-Jacques B, Ma W. Предпочтительный путь рециклинга интернализованным рецептором PGE2 EP4 после стимуляции агонистом в культивируемых нейронах ганглиев дорзального корешка способствует повышенной чувствительности рецептора EP4.Неврология. 2016; 326: 56–68.
CAS PubMed Статья Google Scholar
St-Jacques B, Ma W. Периферический простагландин E2 продлевает сенсибилизацию ноцицептивных нейронов ганглиев задних корешков, возможно, облегчая синтез и антероградный аксональный перенос рецепторов EP4. Exp Neurol. 2014; 261: 354–66.
CAS PubMed Статья Google Scholar
Ng KY, Wong YH, Wise H. Глиальные клетки, выделенные из ганглиев задних корешков, экспрессируют рецепторы простагландина E (2) (EP (4)) и простациклина (IP). Eur J Pharmacol. 2011; 661: 42–8.
CAS PubMed Статья Google Scholar
Дженкинс Д.В., Фенюк В., Хамфри П.П. Характеристика типов простаноидных рецепторов, участвующих в опосредовании высвобождения пептидов, связанных с геном кальцитонина, из культивируемых нейронов тройничного нерва крысы. Br J Pharmacol.2001; 134: 1296–302.
CAS PubMed PubMed Central Статья Google Scholar
Ebersberger A, Natura G, Eitner A, Halbhuber KJ, Rost R, Schaible HG. Влияние простагландина D2 на устойчивые к тетродотоксину Na + токи в нейронах DRG взрослой крысы. Боль. 2011; 152: 1114–26.
CAS PubMed Статья Google Scholar
Секероглу А., Якобсен Дж. М., Янсен-Олесен И., Гупта С., Шейхзаде М., Олесен Дж., Бхатт Д. К..Влияние PGD2 на среднюю менингеальную артерию и профиль экспрессии мРНК L-PGD2-синтазы и рецепторов DP в тройнично-сосудистой системе и других структурах обработки боли в головном мозге крыс. Pharmacol Rep., 2017; 69: 50–6.
CAS PubMed Статья Google Scholar
Isensee J, Wenzel C, Buschow R, Weissmann R, Kuss AW, Hucho T. Транскриптомика исключения подгрупп определяет сигнальные белки, которые определяют подклассы TRPV1-положительных нейронов и новую паракринную цепь.PLoS One. 2014; 9: e115731.
PubMed PubMed Central Статья CAS Google Scholar
Нагира Ю., Гото К., Танака Х., Аоки М., Фуру С., Инагаки Н., Томита Ю., Шичиджо М. Простагландин D2 модулирует возбуждение нейронов тройничного ганглия, усиливая аллергический ринит у морских свинок. J Pharmacol Exp Ther. 2016; 357: 273–80.
CAS PubMed Статья Google Scholar
Telleria-Diaz A, Ebersberger A, Vasquez E, Schache F, Kahlenbach J, Schaible HG. Различные эффекты спинномозгового применения простагландина D2 на ответы нейронов дорсального рога с воздействием коленного сустава у нормальных крыс и у крыс с острым воспалением коленного сустава. Неврология. 2008; 156: 184–92.
CAS PubMed Статья Google Scholar
Кольридж Х.М., Кольридж Дж. К., Гинзель К. Х., Бейкер Д. Дж., Банцетт РБ, Моррисон Массачусетс. Стимуляция «раздражающих» рецепторов и афферентных С-волокон в легких простагландинами.Природа. 1976; 264: 451–3.
CAS PubMed Статья Google Scholar
Фаулер Дж. К., Грин Р., Вайнрайх Д. Два кальций-чувствительных всплеска пост-гиперполяризации в висцеральных сенсорных нейронах кролика. J Physiol. 1985; 365: 59–75.
CAS PubMed PubMed Central Статья Google Scholar
Weinreich D, Wonderlin WF. Подавление кальций-зависимых спайков после гиперполяризации увеличивает возбудимость висцеральных сенсорных нейронов кролика.J Physiol. 1987. 394: 415–27.
CAS PubMed PubMed Central Статья Google Scholar
Fowler JC, Wonderlin WF, Weinreich D. Простагландины блокируют Ca2 + -зависимый медленный всплеск после гиперполяризации независимо от эффектов на приток Ca2 + в висцеральных афферентных нейронах. Brain Res. 1985; 345: 345–9.
CAS PubMed Статья Google Scholar
Gold MS, Шустер MJ, Levine JD. Роль Ca (2 +) — зависимой медленной постгиперполяризации в индуцированной простагландином E2 сенсибилизации культивируемых сенсорных нейронов крыс. Neurosci Lett. 1996; 205: 161–4.
CAS PubMed Статья Google Scholar
England S, Bevan S, Docherty RJ. PGE2 модулирует тетродотоксин-резистентный натриевой ток в нейронах ганглия задних корешков новорожденных крыс через каскад циклической АМФ-протеинкиназы a.J Physiol. 1996; 495 (Pt 2): 429–40.
CAS PubMed PubMed Central Статья Google Scholar
Фицджеральд Э.М., Окусе К., Вуд Дж. Н., Дельфин А.С., Мосс С.Дж. цАМФ-зависимое фосфорилирование резистентного к тетродотоксину потенциал-зависимого натриевого канала SNS. J Physiol. 1999; 516 (Pt 2): 433–46.
CAS PubMed PubMed Central Статья Google Scholar
Бейкер М.Д., Чандра С.Ю., Дин И, Ваксман С.Г., Вуд Дж. GTP-индуцированный устойчивый к тетродотоксину Na + ток регулирует возбудимость сенсорных нейронов малого диаметра мыши и крысы. J Physiol. 2003; 548: 373–82.
CAS PubMed PubMed Central Статья Google Scholar
Saab CY, Cummins TR, Waxman SG. GTP gamma S увеличивает ток Nav1.8 в нейронах ганглиев задних корешков малого диаметра. Exp Brain Res. 2003; 152: 415–9.
PubMed Статья Google Scholar
Раш А.М., Ваксман С.Г. PGE2 увеличивает резистентный к тетродотоксину натрий Nav1.9 в нейронах DRG мыши через G-белки. Brain Res. 2004; 1023: 264–71.
CAS PubMed Статья Google Scholar
Gold MS. Натриевые каналы и обезболивающая. Curr Opin Anaesthesiol. 2000; 13: 565–72.
CAS PubMed Статья Google Scholar
Tripathi PK, Cardenas CG, Cardenas CA, Scroggs RS. Повышение регуляции тетродотоксин-чувствительных натриевых токов простагландином Е (2) в ганглиозных клетках дорсального корешка крысы типа 4. Неврология. 2011; 185: 14–26.
CAS PubMed PubMed Central Статья Google Scholar
Хасар С.Г., Gold MS, Левин Дж.Д. Устойчивый к тетродотоксинам натриевый ток опосредует воспалительную боль у крыс. Neurosci Lett. 1998; 256: 17–20.
CAS PubMed Статья Google Scholar
Zhang P, Gan YH. Простагландин E2, активирующий тригеминальный ганглионарный натриевый канал 1.7, вовлекающий воспалительную боль в височно-нижнечелюстном суставе у крыс. Воспаление. 2017; 40: 1102–9.
CAS PubMed Статья Google Scholar
Liu C, Li Q, Su Y, Bao L. Простагландин E2 способствует передаче Na1.8 через его внутриклеточный мотив RRR через протеинкиназный путь a. Движение. 2010; 11: 405–17.
CAS PubMed Статья Google Scholar
Керр Б.Дж., Суслова В., МакМахон С.Б., Вуд Дж. Роль ТТХ-резистентного натриевого канала Nav 1.8 в индуцированной NGF гипералгезии, но не в нейропатической боли. Нейроотчет. 2001; 12: 3077–80.
CAS PubMed Статья Google Scholar
Sekiguchi F, Aoki Y, Nakagawa M, Kanaoka D, Nishimoto Y, Tsubota-Matsunami M, Yamanaka R, Yoshida S, Kawabata A. AKAP-зависимая сенсибилизация Ca (v) 3.2 каналов через EP ( 4) путь рецептор / цАМФ опосредует индуцированную PGE (2) механическую гипералгезию.Br J Pharmacol. 2013; 168: 734–45.
PubMed PubMed Central Статья Google Scholar
Линли Дж. Э., Роуз К., Оои Л., Гампер Н. Понимание воспалительной боли: ионные каналы, способствующие острой и хронической ноцицепции. Pflugers Arch. 2010. 459: 657–69.
CAS PubMed Статья Google Scholar
Гамильтон С.Г., Уэйд А., МакМахон С.Б.Влияние воспаления и медиаторов воспаления на ноцицептивное поведение, индуцированное аналогами АТФ у крыс. Br J Pharmacol. 1999; 126: 326–32.
CAS PubMed PubMed Central Статья Google Scholar
Ван Ц., Ли Г.В., Хуанг Л.Й. Простагландин E2 потенцирование опосредованных рецептором P2X3 токов в нейронах ганглиев задних корешков. Молочная боль. 2007; 3:22.
PubMed PubMed Central Статья CAS Google Scholar
Ван Ц., Гу И, Ли GW, Хуан Л. Критическая роль датчика цАМФ Epac в переключении передачи сигналов протеинкиназы в индуцированном простагландином E2 потенцировании токов рецептора P2X3 у воспаленных крыс. J Physiol. 2007; 584: 191–203.
CAS PubMed PubMed Central Статья Google Scholar
Gu Y, Wang C, Li G, Huang LY. ЭКСПРЕСС: F-актин связывает передачу сигналов Epac-PKC с сенсибилизацией пуринергических рецепторов P2X3 в ганглиях задних корешков после воспаления.Молочная боль. 2016; 12.
Артикул CAS Google Scholar
Prado FC, Araldi D, Vieira AS, Oliveira-Fusaro MC, Tambeli CH, Parada CA. Активация нейронального рецептора P2X3 необходима для гипералгезии, вызванной простагландинами и симпатомиметическими аминами, высвобождаемыми во время воспаления. Нейрофармакология. 2013; 67: 252–8.
CAS PubMed Статья Google Scholar
Kim BM, Lee SH, Shim WS, Oh U. Гистамин-индуцированный приток Ca (2+) через путь PLA (2) / липоксигеназа / TRPV1 в сенсорных нейронах крыс. Neurosci Lett. 2004. 361: 159–62.
CAS PubMed Статья Google Scholar
Shim WS, Tak MH, Lee MH, Kim M, Koo JY, Lee CH, Oh U. TRPV1 опосредует вызванный гистамином зуд посредством активации фосфолипазы A2 и 12-липоксигеназы. J Neurosci. 2007. 27: 2331–7.
CAS PubMed PubMed Central Статья Google Scholar
Hwang SW, Oh U. Современные концепции ноцицепции: ноцицептивные молекулярные сенсоры в сенсорных нейронах. Curr Opin Anaesthesiol. 2007. 20: 427–34.
PubMed Статья Google Scholar
Choi SI, Lim JY, Yoo S, Kim H, Hwang SW. Возникающая роль TRPV1 спинного мозга в обострении боли. Neural Plast. 2016; 2016: 5954890.
PubMed PubMed Central Статья CAS Google Scholar
Ким Й.Х., Бэк С.К., Дэвис А.Дж., Чжон Х., Джо Х.Дж., Чанг Джи, На ХС, Бэ Ю.С., Ким С.Дж., Ким Дж.С. TRPV1 в ГАМКергических интернейронах опосредует нейропатическую механическую аллодинию и растормаживание ноцицептивных цепей в спинном мозге. Нейрон. 2012; 74: 640–7.
CAS PubMed Статья Google Scholar
Ма В., Квирион Р. Медиаторы воспаления, модулирующие временный рецепторный потенциал ваниллоидного 1 рецептора: терапевтические мишени для лечения воспалительной и невропатической боли.Эксперт считает, что цели. 2007; 11: 307–20.
PubMed Статья Google Scholar
Pitchford S, Levine JD. Простагландины сенсибилизируют ноцицепторы в культуре клеток. Neurosci Lett. 1991; 132: 105–8.
CAS PubMed Статья Google Scholar
Lopshire JC, Nicol GD. Каскад трансдукции цАМФ опосредует простагландин E2 усиление вызванного капсаицином тока в сенсорных нейронах крыс: исследования на всех клетках и одноканальные исследования.J Neurosci. 1998; 18: 6081–92.
CAS PubMed PubMed Central Статья Google Scholar
Морияма Т., Хигаши Т., Тогаши К., Иида Т., Сеги Е., Сугимото Ю., Томинага Т., Нарумия С., Томинага М. Сенсибилизация TRPV1 с помощью EP1 и IP выявляет периферический ноцицептивный механизм простагландинов. Молочная боль. 2005; 1: 3.
PubMed PubMed Central Статья CAS Google Scholar
Намба Т., Оида Н., Сугимото Ю., Какидзука А., Негиси М., Итикава А., Нарумия С. Клонирование кДНК рецептора простациклина мыши. Множественные сигнальные пути и экспрессия в мозговом веществе тимуса. J Biol Chem. 1994; 269: 9986–92.
CAS PubMed Google Scholar
Шницлер К., Шутов Л.П., Ван Канеган М.Дж., Меррилл М.А., Николс Б., Макнайт Г.С., Страк С., Ад Дж. В., Усачев Ю.М.. Заякоривание протеинкиназы a через AKAP150 необходимо для модуляции TRPV1 форсколином и простагландином E2 в сенсорных нейронах мыши.J Neurosci. 2008; 28: 4904–17.
CAS PubMed PubMed Central Статья Google Scholar
Йеске Н.А., Диоген А., Рупарел Н.Б., Ференбахер Дж. К., Генри М., Акопян А. Н., Харгривз К. М.. Заякоренный белок А-киназы опосредует тепловую гипералгезию TRPV1 через PKA-фосфорилирование TRPV1. Боль. 2008. 138: 604–16.
CAS PubMed PubMed Central Статья Google Scholar
Ma W, St-Jacques B, Rudakou U, Kim YN. Стимуляция экстернализации и синтеза TRPV1 в нейронах ганглия задних корешков способствует усилению активности TRPV1 PGE2 и сенсибилизации ноцицепторов. Eur J Pain. 2017; 21: 575–93.
CAS PubMed Статья Google Scholar
Алессандри-Хабер Н., Йе Дж. Дж., Бойд А. Э., Парада, Калифорния, Чен Х, Рейхлинг Д. Б., Левин Дж. Д.. Гипотоничность вызывает у крыс опосредованную TRPV4 ноцицепцию.Нейрон. 2003. 39: 497–511.
CAS PubMed Статья Google Scholar
Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Транзиторный рецепторный потенциал ваниллоида 4 необходим при нейропатической боли, вызванной химиотерапией, у крыс. J Neurosci. 2004. 24: 4444–52.
CAS PubMed PubMed Central Статья Google Scholar
Алессандри-Хабер Н., Джозеф Э., Дина О.А., Лидтке В., Левин Дж. Д.. TRPV4 опосредует поведение, связанное с болью, вызванное легкими гипертоническими стимулами в присутствии медиатора воспаления. Боль. 2005; 118: 70–9.
CAS PubMed Статья Google Scholar
Алессандри-Хабер Н., Дина О.А., Джозеф Е.К., Райхлинг Д., Левин Д.Д. Переходный рецепторный потенциал ваниллоид-4-зависимого механизма гипералгезии задействован согласованным действием медиаторов воспаления.J Neurosci. 2006; 26: 3864–74.
CAS PubMed PubMed Central Статья Google Scholar
Chen X, Alessandri-Haber N, Levine JD. Заметное ослабление индуцированной медиатором воспаления сенсибилизации С-волокон к механическим и гипотоническим стимулам у мышей TRPV4 — / -. Молочная боль. 2007; 3:31.
PubMed PubMed Central Google Scholar
Li CL, Li KC, Wu D, Chen Y, Luo H, Zhao JR, Wang SS, Sun MM, Lu YJ, Zhong YQ и др.Типы соматосенсорных нейронов, идентифицированные с помощью секвенирования одноклеточной РНК с высоким охватом и функциональной гетерогенности. Cell Res. 2016; 26: 83–102.
CAS PubMed Статья PubMed Central Google Scholar
Ю С., Чхве С.И., Ли С., Сон Дж., Ян Си, Банг С., Ким Су, Мин К.Х., Хван СВ. Эндогенная экспрессия TRPV4 гибридной нейрональной клеточной линии N18D3 и ее использование для поиска нового синтетического лиганда. J Mol Neurosci. 2017; 63: 422–30.
CAS PubMed Статья PubMed Central Google Scholar
Bang S, Yang TJ, Yoo S, Heo TH, Hwang SW. Ингибирование сенсорных нейрональных TRP способствует антиноцицепции бутамбена. Neurosci Lett. 2012; 506: 297–302.
CAS PubMed Статья PubMed Central Google Scholar
Bang S, Yoo S, Yang TJ, Cho H, Hwang SW.Ноцицептивные и провоспалительные эффекты диметилаллилпирофосфата через активацию TRPV4. Br J Pharmacol. 2012; 166: 1433–43.
CAS PubMed PubMed Central Статья Google Scholar
Bang S, Yoo S, Yang TJ, Cho H, Kim YG, Hwang SW. Резолвин D1 ослабляет активацию сенсорных временных каналов потенциальных рецепторов, что приводит к множественной антиноцицепции. Br J Pharmacol. 2010; 161: 707–20.
CAS PubMed PubMed Central Статья Google Scholar
Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, Earley TJ, Patapoutian A. Ядовитый канал холодных ионов TRPA1 активируется едкими соединениями и брадикинином. Нейрон. 2004. 41: 849–57.
CAS PubMed Статья Google Scholar
Kim S, Hwang SW. Новые роли TRPA1 в ощущении окислительного стресса и его значение в защите и опасности. Arch Pharm Res. 2013; 36: 783–91.
CAS PubMed Статья Google Scholar
Dall’Acqua MC, Bonet IJ, Zampronio AR, Tambeli CH, Parada CA, Fischer L. Вклад временного рецепторного потенциала анкирина 1 (TRPA1) в ноцицептивные эффекты простагландина E in vivo (2). Life Sci. 2014; 105: 7–13.
CAS PubMed Статья Google Scholar
Чо Х, Ян Й.Д., Ли Дж., Ли Б., Ким Т., Чан И, Бэк С.К., На ХС, Харфе Б.Д., Ван Ф. и др. Аноктамин 1 хлоридного канала, активируемый кальцием, действует как датчик тепла в ноцицептивных нейронах.Nat Neurosci. 2012; 15: 1015–21.
CAS PubMed Статья Google Scholar
Моррис М.Э., Ди Костанцо Г.А., Фокс С., Верман Р. Деполяризующее действие ГАМК (гамма-аминомасляная кислота) на миелинизированные волокна периферических нервов. Brain Res. 1983; 278: 117–26.
CAS PubMed Статья PubMed Central Google Scholar
Funk K, Woitecki A, Franjic-Wurtz C., Gensch T., Mohrlen F, Frings S.Модуляция хлоридного гомеостаза медиаторами воспаления в нейронах ганглия задних корешков. Молочная боль. 2008; 4:32.
PubMed PubMed Central Статья CAS Google Scholar
Усоскин Д., Фурлан А., Ислам С., Абдо Х., Лоннерберг П., Лу Д., Хьерлинг-Леффлер Дж., Хеггстром Дж., Харченко О., Харченко П.В. и др. Беспристрастная классификация типов сенсорных нейронов путем крупномасштабного секвенирования одноклеточной РНК. Nat Neurosci.2015; 18: 145–53.
CAS PubMed Статья PubMed Central Google Scholar
Мао С., Гарсон-Мувди Т., Ди Фульвио М., Чен Й., Дельпир Е., Альварес Ф.Дж., Альварес-Лифманс Ф.Дж. Молекулярная и функциональная экспрессия котранспортеров хлорида катионов в нейронах ганглия дорсального корешка во время постнатального созревания. J Neurophysiol. 2012; 108: 834–52.
PubMed PubMed Central Статья Google Scholar
Gilbert D, Franjic-Wurtz C, Funk K, Gensch T., Frings S, Mohrlen F. Дифференциальное созревание гомеостаза хлоридов в первичных афферентных нейронах соматосенсорной системы. Int J Dev Neurosci. 2007. 25: 479–89.
CAS PubMed Статья Google Scholar
Моралес-Аза Б.М., Чиллингворт Н.Л., Пейн Дж. А., Дональдсон Л. Ф.. Воспаление изменяет экспрессию котранспортера катионхлорида в сенсорных нейронах. Neurobiol Dis. 2004; 17: 62–9.
CAS PubMed Статья Google Scholar
Канака С., Оно К., Окабе А., Курияма К., Ито Т., Фукуда А., Сато К. Характер дифференциальной экспрессии матричных РНК, кодирующих котранспортеры K-cl (KCC1,2) и Na-K-2Cl котранспортер (NKCC1) в нервной системе крыс. Неврология. 2001; 104: 933–46.
CAS PubMed Статья Google Scholar
Лу Дж., Карадшех М., Дельпир Е. Регуляция развития нейронально-специфической изоформы котранспортера K-cl KCC2 в постнатальном мозге крыс. J Neurobiol. 1999; 39: 558–68.
CAS PubMed Статья Google Scholar
Эмери Э., Янг Г.Т., Макнотон, Пенсильвания. Ионные каналы HCN2: растущая роль стимуляторов боли. Trends Pharmacol Sci. 2012; 33: 456–63.
CAS PubMed Статья Google Scholar
Chaplan SR, Guo HQ, Lee DH, Luo L, Liu C, Kuei C, Velumian AA, Butler MP, Brown SM, Dubin AE. Каналы кардиостимулятора, активируемые гиперполяризацией нейронов, вызывают невропатическую боль. J Neurosci. 2003. 23: 1169–78.
CAS PubMed PubMed Central Статья Google Scholar
Tu H, Deng L, Sun Q, Yao L, Han JS, Wan Y. Активируемые гиперполяризацией циклические нуклеотидно-зависимые катионные каналы: роли в дифференциальных электрофизиологических свойствах первичных афферентных нейронов крысы.J Neurosci Res. 2004. 76: 713–22.
CAS PubMed Статья Google Scholar
Момин А., Кадиу Х., Мейсон А., Макнотон, Пенсильвания. Роль тока Ih, активируемого гиперполяризацией, в соматосенсорных нейронах. J Physiol. 2008; 586: 5911–29.
CAS PubMed PubMed Central Статья Google Scholar
Чо Х. Дж., Стайкопулос В., Фернесс Дж. Б., Дженнингс Э. А..Вызванное воспалением увеличение активируемого гиперполяризацией, циклического нуклеотид-зависимого белка канала в нейронах тройничного ганглия и эффект бупренорфина. Неврология. 2009. 162: 453–61.
CAS PubMed Статья Google Scholar
Ingram SL, Williams JT. Опиоидное ингибирование Ih с помощью аденилатциклазы. Нейрон. 1994; 13: 179–86.
CAS PubMed Статья Google Scholar
Ingram SL, Уильямс JT. Модуляция тока, активируемого гиперполяризацией (Ih), циклическими нуклеотидами в первичных афферентных нейронах морских свинок. J Physiol. 1996; 492 (Pt 1): 97–106.
CAS PubMed PubMed Central Статья Google Scholar
Эмери EC, Young GT, Berrocoso EM, Chen L, McNaughton PA. Ионные каналы HCN2 играют центральную роль в воспалительной и невропатической боли. Наука. 2011; 333: 1462–6.
CAS PubMed Статья Google Scholar
Хан Г.М., Чен С.Р., Пан Х.Л. Роль первичных афферентных нервов в аллодинии, вызванной диабетической невропатией у крыс. Неврология. 2002; 114: 291–9.
CAS PubMed Статья Google Scholar
Сон Й, Ли Х.М., Се Р.Г., Юэ З.Ф., Сонг ХДЖ, Ху SJ, Син JL. Вызванный разрыв в поврежденных нейронах ганглия дорзального корешка Abeta: механизм, лежащий в основе тактильной аллодинии. Боль. 2012; 153: 657–65.
PubMed Статья Google Scholar
Сяо WH, Беннетт Дж. Стойкие низкочастотные спонтанные разряды в первичных афферентных нейронах А-волокон и С-волокон во время воспалительного болевого состояния. Анестезиология. 2007; 107: 813–21.
CAS PubMed Статья Google Scholar
Дуан Б., Ченг Л., Буран С., Бритц О, Падилья С., Гарсия-Кампмани Л., Крашес М., Ноултон В., Веласкес Т., Рен Х и др. Определение контуров позвоночника, передающих и блокирующих механическую боль.Клетка. 2014; 159: 1417–32.
CAS PubMed PubMed Central Статья Google Scholar
Ван Х., Энерт К., Бреннер Г.Дж., Вульф С.Дж. Брадикинин и периферическая сенсибилизация. Biol Chem. 2006; 387: 11–4.
CAS PubMed Google Scholar
Выклицкий Л., Кноткова-Урбанцова Н., Витаскова З., Влахова В., Кресс М., Рих П.В. Медиаторы воспаления при кислом pH активируют рецепторы капсаицина в культивируемых сенсорных нейронах новорожденных крыс.J Neurophysiol. 1998. 79: 670–6.
CAS PubMed Статья Google Scholar
Смит Дж. А., Дэвис К. Л., Берджесс Г. М.. Индуцированная простагландином E2 сенсибилизация вызванных брадикинином ответов в нейронах ганглия задних корешков крыс опосредуется цАМФ-зависимой протеинкиназой а. Eur J Neurosci. 2000; 12: 3250–8.
CAS PubMed Статья Google Scholar
Касаи М., Мизумура К. Эффекты PGE (2) на нейронах ганглиев задних корешков крыс у интактных и воспаленных адъювантом крыс: роль NGF в деполяризации, вызванной PGE (2). Neurosci Res. 2001; 41: 345–53.
CAS PubMed Статья Google Scholar
Gold MS, Gebhart GF. Сенсибилизация ноцицепторов в патогенезе боли. Nat Med. 2010; 16: 1248–57.
CAS PubMed PubMed Central Статья Google Scholar
Taiwo YO, Levine JD. Характеристика метаболитов арахидоновой кислоты, опосредующих брадикинин и гипералгезию норадреналина. Brain Res. 1988. 458: 402–6.
CAS PubMed Статья Google Scholar
Cunha TM, Verri WA, Fukada SY, Guerrero AT, Santodomingo-Garzón T, Poole S, Parada CA, Ferreira SH, Cunha FQ. TNF-α и IL-1β опосредуют воспалительную гиперноцицепцию у мышей, запускаемую B 1, но не B 2 кининовым рецептором.Eur J Pharmacol. 2007; 573: 221–9.
CAS PubMed Статья Google Scholar
Феррейра С., Лоренцетти Б., Кунха Ф., Пул С. Высвобождение брадикинина TNF-α играет ключевую роль в развитии воспалительной гипералгезии. Inflamm Res. 1993; 38: C7–9.
CAS Google Scholar
Пул С., Лоренцетти Б., Кунха Дж., Кунья Ф., Феррейра С. Рецепторы брадикинина B1 и B2, фактор некроза опухоли α и воспалительная гипералгезия.Br J Pharmacol. 1999; 126: 649–56.
CAS PubMed PubMed Central Статья Google Scholar
Oshita K, Inoue A, Tang HB, Nakata Y, Kawamoto M, Yuge O. Стимуляция каннабиноидного рецептора CB1 модулирует временную активность рецептора ваниллоидного рецептора 1 в притоке кальция и высвобождении вещества P в культивируемых ганглиозных клетках дорсального корешка крысы . J Pharmacol Sci. 2005. 97: 377–85.
CAS PubMed Статья Google Scholar
Kopp UC, Cicha MZ, Nakamura K, Nusing RM, Smith LA, Hokfelt T. Активация рецепторов EP4 способствует простагландин E2-опосредованной стимуляции почечных сенсорных нервов. Am J Physiol Renal Physiol. 2004; 287: F1269–82.
CAS PubMed Статья Google Scholar
Segond von Banchet G, Scholze A, Schaible HG. Простагландин E2 увеличивает экспрессию рецептора нейрокинина 1 во взрослых сенсорных нейронах в культуре: новая роль простагландинов.Br J Pharmacol. 2003. 139: 672–80.
CAS PubMed Статья Google Scholar
Chiu IM, von Hehn CA, Woolf CJ. Нейрогенное воспаление и периферическая нервная система в защите хозяина и иммунопатологии. Nat Neurosci. 2012; 15: 1063–7.
CAS PubMed PubMed Central Статья Google Scholar
Энтони Д., Ясинский Д.М. Послеоперационное обезболивание: морфин по сравнению с кеторолаком.J Perianesth Nurs. 2002; 17: 30–42.
PubMed Статья Google Scholar
Тумати С., Роеске В.Р., Вандера Т.В., Варга Е.В. Продолжительное лечение морфином увеличивает вызванное простагландином E2 высвобождение пептида, связанного с геном кальцитонина, из первичных сенсорных нейронов зависимым от PKA образом. Eur J Pharmacol. 2010; 648: 95–101.
CAS PubMed PubMed Central Статья Google Scholar
Томпсон SW, Беннетт Д.Л., Керр Б.Дж., Брэдбери Е.Дж., МакМахон С.Б. Нейротрофический фактор головного мозга является эндогенным модулятором ноцицептивных ответов в спинном мозге. Proc Natl Acad Sci U S. A. 1999; 96: 7714–8.
CAS PubMed PubMed Central Статья Google Scholar
Чжоу XF, Chie ET, Deng YS, Zhong JH, Xue Q, Rush RA, Xian CJ. Поврежденные первичные сенсорные нейроны переключают фенотип на нейротрофический фактор головного мозга крысы.Неврология. 1999; 92: 841–53.
CAS PubMed Статья Google Scholar
Мериги А., Салио С., Гирри А., Лосси Л., Феррини Ф., Бетелли С., Бардони Р. BDNF как модулятор боли. Prog Neurobiol. 2008. 85: 297–317.
CAS PubMed Статья Google Scholar
Ван Х, Ратнам Дж., Зоу Б., Англия П.М., Басбаум А.И. Передача сигналов TrkB необходима как для индукции, так и для поддержания постоянной боли, вызванной повреждением тканей и нервов.J Neurosci. 2009; 29: 5508–15.
CAS PubMed PubMed Central Статья Google Scholar
Quintao NL, Santos AR, Campos MM, Calixto JB. Роль нейротрофических факторов в возникновении и поддержании механической гиперноцицепции после отрыва плечевого сплетения у мышей. Боль. 2008. 136: 125–33.
CAS PubMed Статья Google Scholar
Choi SY, Choi BH, Suh BC, Chae HD, Kim JS, Shin MJ, Kang SS, Negishi M, Kim KT. Усиление опосредованной PGE (2) продукции цАМФ во время нейрональной дифференцировки C-клеток нейробластомы человека SK-N-BE (2). J Neurochem. 2001; 79: 303–10.
CAS PubMed Статья Google Scholar
Ким Х.С., Сон М., Ким И, Рю Ш, Сух П.Г. Дексаметазон дифференцирует клетки NG108-15 за счет индукции циклооксигеназы 1. Exp Mol Med.2003; 35: 203–10.
CAS PubMed Статья Google Scholar
Mitani K, Sekiguchi F, Maeda T., Tanaka Y, Yoshida S, Kawabata A. Путь рецептора простагландина E2 / EP4 / циклического AMP / Ca (2+) канала T-типа опосредует нейритогенез в сенсорных нейронах. как клетки ND7 / 23. J Pharmacol Sci. 2016; 130: 177–80.
CAS PubMed Статья Google Scholar
Nango H, Kosuge Y, Miyagishi H, Sugawa K, Ito Y, Ishige K. Простагландин E2 способствует росту нейритов в клеточной линии, подобной двигательным нейронам, NSC-34. J Pharmacol Sci. 2017; 135: 64–71.
CAS PubMed Статья Google Scholar
Hiruma H, Ichikawa T, Kobayashi H, Hoka S, Takenaka T, Kawakami T. Простагландин E (2) усиливает аксональный транспорт и нейритогенез в культивируемых нейронах ганглиев дорсальных корешков мыши. Неврология.2000; 100: 885–91.
CAS PubMed Статья Google Scholar
Sondell M, Lundborg G, Kanje M. Фактор роста эндотелия сосудов обладает нейротрофической активностью и стимулирует рост аксонов, увеличивая выживаемость клеток и пролиферацию шванновских клеток в периферической нервной системе. J Neurosci. 1999; 19: 5731–40.
CAS PubMed PubMed Central Статья Google Scholar
Kelton JG, Blajchman MA. Простагландин I2 (простациклин). Кан Мед Асс Дж. 1980; 122: 175–9.
CAS PubMed PubMed Central Google Scholar
Taiwo YO, Bjerknes LK, Goetzl EJ, Levine JD. Посредничество первичной афферентной периферической гипералгезии с помощью системы вторичных мессенджеров цАМФ. Неврология. 1989; 32: 577–80.
CAS PubMed Статья Google Scholar
Омана-Сапата I, Блей КР. Стабильный аналог простациклина усиливает эктопическую активность сенсорных нейронов крыс после невропатического повреждения. Brain Res. 2001; 904: 85–92.
CAS PubMed Статья Google Scholar
Накаэ К., Хаяси Ф., Хаяси М., Ямамото Н., Иино Т., Йошикава С., Гупта Дж. Функциональная роль рецептора простациклина в нейронах ганглиев задних корешков крыс. Neurosci Lett. 2005; 388: 132–7.
CAS PubMed Статья Google Scholar
Best LC, Мартин Т.Дж., Рассел Р.Г., Престон FE. Простациклин увеличивает уровни циклического АМФ и активность аденилатциклазы в тромбоцитах. Природа. 1977; 267: 850–2.
CAS PubMed Статья Google Scholar
Мурата Т., Ушикуби Ф, Мацуока Т., Хирата М., Ямасаки А., Сугимото И., Итикава А., Азе И., Танака Т., Йошида Н. и др. Изменение восприятия боли и воспалительной реакции у мышей, лишенных рецептора простациклина. Природа.1997; 388: 678–82.
CAS PubMed Статья Google Scholar
Nakae K, Saito K, Iino T, Yamamoto N, Wakabayashi M, Yoshikawa S, Matsushima S, Miyashita H, Sugimoto H, Kiba A, Gupta J. Антагонист рецепторов простациклина подавляет сенсибилизированное высвобождение вещества P. из сенсорных нейронов крысы. J Pharmacol Exp Ther. 2005; 315: 1136–42.
CAS PubMed Статья Google Scholar
Андерссон Д.А., Джентри С., Мосс С., Беван С. Переходный рецепторный потенциал A1 является сенсорным рецептором для множества продуктов окислительного стресса. J Neurosci. 2008. 28: 2485–94.
CAS PubMed PubMed Central Статья Google Scholar
Materazzi S, Nassini R, Andre E, Campi B, Amadesi S, Trevisani M, Bunnett NW, Patacchini R, Geppetti P. Кокс-зависимые метаболиты жирных кислот вызывают боль через активацию раздражающего рецептора TRPA1.Proc Natl Acad Sci U S. A. 2008; 105: 12045–50.
CAS PubMed PubMed Central Статья Google Scholar
Крус-Оренго Л., Дакка А., Хойерманн Р.Дж., Янг Т.Дж., Монтана М.С., Кавано Е.Дж., Ким Д., Стори Г.М. Кожная ноцицепция, вызванная 15-дельта PGJ2 через активацию ионного канала TRPA1. Молочная боль. 2008; 4:30.
PubMed PubMed Central Статья CAS Google Scholar
Weng Y, Batista-Schepman PA, Barabas ME, Harris EQ, Dinsmore TB, Kossyreva EA, Foshage AM, Wang MH, Schwab MJ, Wang VM, et al. Метаболит простагландина вызывает ингибирование TRPA1 и канально-зависимую ноцицепцию. Молочная боль. 2012; 8: 75.
CAS PubMed PubMed Central Статья Google Scholar
Чури С.Б., Абдель-Алим О.С., Тамбер К.К., Скудери-Портер Х., Тейлор Б.К. Розиглитазон интратекально действует на рецептор-гамма, активируемый пролифератором пероксисом, для быстрого подавления невропатической боли у крыс.J Pain. 2008; 9: 639–49.
CAS PubMed PubMed Central Статья Google Scholar
Napimoga MH, Souza GR, Cunha TM, Ferrari LF, Clemente-Napimoga JT, Parada CA, Verri WA Jr, Cunha FQ, Ferreira SH. 15d-простагландин J2 подавляет воспалительную гиперноцицепцию: вовлечение периферических опиоидных рецепторов. J Pharmacol Exp Ther. 2008. 324: 313–21.
CAS PubMed Статья Google Scholar
Pena-dos-Santos DR, Severino FP, Pereira SA, Rodrigues DB, Cunha FQ, Vieira SM, Napimoga MH, Clemente-Napimoga JT. Активация периферических каппа / дельта опиоидных рецепторов опосредует индуцированную 15-дезокси- (Delta12,14) -простагландином J2 антиноцицепцию в височно-нижнечелюстном суставе крысы. Неврология. 2009; 163: 1211–9.
CAS PubMed Статья Google Scholar
Hinman A, Chuang HH, Bautista DM, Julius D. Активация канала TRP путем обратимой ковалентной модификации.Proc Natl Acad Sci U S. A. 2006; 103: 19564–8.
CAS PubMed PubMed Central Статья Google Scholar
Bang S, Hwang SW. Чувствительность TRPA1 к полимодальным лигандам и способы его взаимодействия. J Gen Physiol. 2009. 133: 257–62.
CAS PubMed PubMed Central Статья Google Scholar
Макферсон Л.Дж., Дубин А.Е., Эванс М.Дж., Марр Ф., Шульц П.Г., Краватт Б.Ф., Патапутян А.Вредные соединения активируют ионные каналы TRPA1 посредством ковалентной модификации цистеинов. Природа. 2007; 445: 541–5.
CAS PubMed Статья Google Scholar
Takahashi N, Mizuno Y, Kozai D, Yamamoto S, Kiyonaka S, Shibata T., Uchida K, Mori Y. Молекулярная характеристика активации канала TRPA1 цистеин-реактивными медиаторами воспаления. Каналы (Остин). 2008; 2: 287–98.
Артикул Google Scholar
Тейлор-Кларк Т.Э., Ундем Б.Дж., Макглашан Д.В. мл., Гхатта С., Карр М.Дж., МакАлександр М.А. Простагландин-индуцированная активация ноцицептивных нейронов посредством прямого взаимодействия с временным рецепторным потенциалом A1 (TRPA1). Mol Pharmacol. 2008. 73: 274–81.
CAS PubMed Статья Google Scholar
Кападиа Р., Йи Дж. Х., Вемуганти Р. Механизмы противовоспалительного и нейрозащитного действия агонистов PPAR-гамма. Front Biosci.2008; 13: 1813–26.
CAS PubMed PubMed Central Статья Google Scholar
Ricote M, Li AC, Willson TM, Kelly CJ, Glass CK. Гамма-рецептор, активируемый пролифератором пероксисом, является негативным регулятором активации макрофагов. Природа. 1998. 391: 79–82.
CAS PubMed Статья Google Scholar
Мукерджи Р., Джоу Л., Нунан Д., Макдоннелл Д.П.Рецепторы, активируемые пролифератором пероксисом (PPAR) человека и крысы, демонстрируют сходное распределение в тканях, но различную чувствительность к активаторам PPAR. J Стероид Biochem Mol Biol. 1994; 51: 157–66.
CAS PubMed Статья Google Scholar
Ягами Т., Ямамото Ю., Кома Х. Физиологические и патологические роли 15-дезокси-дельта (12,14) -простагландина J2 в центральной нервной системе и неврологических заболеваниях. Mol Neurobiol.2018; 55: 2227–48.
CAS PubMed Статья Google Scholar
Хамберг М., Свенссон Дж., Самуэльссон Б. Тромбоксаны: новая группа биологически активных соединений, полученных из эндопероксидов простагландинов. Proc Natl Acad Sci U S. A. 1975; 72: 2994–8.
CAS PubMed PubMed Central Статья Google Scholar
Needleman P, Moncada S, Bunting S, Vane JR, Hamberg M, Samuelsson B.Идентификация фермента в микросомах тромбоцитов, который генерирует тромбоксан А2 из эндопероксидов простагландинов. Природа. 1976; 261: 558–60.
CAS PubMed Статья Google Scholar
Хирата М., Хаяси Ю., Ушикуби Ф., Йокота Ю., Кагеяма Р., Наканиши С., Нарумия С. Клонирование и экспрессия кДНК для рецептора тромбоксана А2 человека. Природа. 1991; 349: 617–20.
CAS PubMed Статья Google Scholar
de Souza BF, Tricoci P. Новые антитромбоцитарные агенты: основное внимание уделяется антагонистам рецепторов тромбина. J Cardiovasc Transl Res. 2013; 6: 415–24.
Артикул Google Scholar
Эллинсворт округ Колумбия, Шукла Н., Флеминг I, Джереми Дж. Взаимодействие между тромбоксаном а (2), рецепторами тромбоксана / простагландина (ТР) и гиперполяризацией эндотелия. Cardiovasc Res. 2014; 102: 9–16.
CAS PubMed Статья Google Scholar
Накахата Н. Тромбоксан A2: физиология / патофизиология, трансдукция клеточного сигнала и фармакология. Pharmacol Ther. 2008. 118: 18–35.
CAS PubMed Статья Google Scholar
Karla W, Shams H, Orr JA, Scheid P. Влияние миметика тромбоксана A2, U46,619, на афференты легочного блуждающего нерва у кошек. Respir Physiol. 1992; 87: 383–96.
CAS PubMed Статья Google Scholar
Шамс Х., Шейд П. Влияние тромбоксана на дыхание и малое кровообращение у кошек: роль блуждающего нерва. J. Appl Physiol (1985). 1990; 68: 2042–6.
CAS Статья Google Scholar
Айзава Х., Хиросе Т. Возможный механизм гиперчувствительности дыхательных путей, вызванный простагландином F2 альфа и тромбоксаном A2. Простагландины Leukot Essent Fatty Acids. 1988; 33: 185–9.
CAS PubMed Google Scholar
Pickar JG. Миметик тромбоксана А2 U-46619 подавляет соматомоторную активность за счет блуждающего рефлекса легкого. Am J Phys. 1998. 275: R706–12.
CAS Google Scholar
Carrithers JA, Liu F, Shirer HW, Orr JA. Механизмы тахипного ответа на миметик тромбоксана А2 U-46 619 у кроликов. Am J Phys. 1994; 266: R321–7.
CAS Google Scholar
Вакер М.Дж., Тегерани Р.Н., Смут Р.Л., Орр Дж.А. Миметик тромбоксана а (2) вызывает брадикардию, опосредованную стимуляцией афферентных нервов блуждающего нерва. Am J Physiol Heart Circ Physiol. 2002; 282: h582–90.
CAS PubMed Статья Google Scholar
Sun SY, Wang W, Schultz HD. Активация сердечных афферентов арахидоновой кислотой: относительный вклад метаболических путей. Am J Physiol Heart Circ Physiol. 2001; 281: H93 – h204.
CAS PubMed Статья Google Scholar
Kenagy J, VanCleave J, Pazdernik L, Orr JA. Стимуляция афферентных нервов III и IV групп от задней конечности тромбоксаном А2. Brain Res. 1997; 744: 175–8.
CAS PubMed Статья Google Scholar
Mense S. Болезненное восприятие скелетными мышцами в связи с клинической болью в мышцах. Боль.1993; 54: 241–89.
CAS PubMed Статья Google Scholar
Mitchell JH, Kaufman MP, Iwamoto GA. Прессорный рефлекс при физической нагрузке: его сердечно-сосудистые эффекты, афферентные механизмы и центральные пути. Annu Rev Physiol. 1983; 45: 229–42.
CAS PubMed Статья Google Scholar
Fu LW, Guo ZL, Longhurst JC. Неизвестная роль эндогенного тромбоксана А2 в активации симпатических афферентов сердца во время ишемии.J Physiol. 2008. 586: 3287–300.
CAS PubMed PubMed Central Статья Google Scholar
Wacker MJ, Tyburski JB, Ammar CP, Adams MC, Orr JA. Обнаружение мРНК рецептора тромбоксана а (2) в нейронах узлового ганглия кролика. Neurosci Lett. 2005; 386: 121–6.
CAS PubMed Статья Google Scholar
Wacker MJ, Tevis O, Hanke J, Howard T., Gilbert W., Orr JA.Характеристика экспрессии рецептора тромбоксана а (2) и мРНК TRPV1 в культивируемых сенсорных нейронах. Neurosci Lett. 2012; 515: 12–7.
CAS PubMed PubMed Central Статья Google Scholar
Андох Т., Нисикава Ю., Ямагути-Миямото Т., Нодзима Х., Нарумия С., Кураиши Ю. Тромбоксан А2 вызывает у мышей реакции, связанные с зудом, через рецепторы TP в коже. J Invest Dermatol. 2007; 127: 2042–7.
CAS PubMed Статья Google Scholar
Мур Р.А., Дерри С., Олдингтон Д., Коул П., Виффен П.Дж. Нежелательные явления, связанные с однократным приемом пероральных анальгетиков при острой послеоперационной боли у взрослых — обзор Кокрановских обзоров. Кокрановская база данных Syst Rev.2015; 10: CD011407.
Езерски Р.П., Ханссон П. Воспалительная и невропатическая боль от скамьи до постели: что пошло не так? J Pain. 2018; 19: 571–88.
PubMed Статья Google Scholar
Lim JY, Park CK, Hwang SW. Биологическая роль резолвина и родственных ему веществ в разрешении боли. Biomed Res Int. 2015; 2015: 830930.
PubMed PubMed Central Google Scholar
Choi G, Hwang SW. Модуляция активности нейронных ионных каналов про-резольвентами, полученными из жирных кислот. Front Physiol. 2016; 7: 523.
PubMed PubMed Central Статья Google Scholar
Джонс Р.Л., Гимбыч М.А., Вудворд Д.Ф. Антагонисты простаноидных рецепторов: стратегии развития и терапевтические применения. Br J Pharmacol. 2009; 158: 104–45.
CAS PubMed PubMed Central Статья Google Scholar
Хирата Т., Нарумия С. Простаноидные рецепторы. Chem Rev.2011; 111: 6209–30.
CAS PubMed Статья Google Scholar
Ott D, Simm B, Pollatzek E, Gerstberger R, Rummel C, Roth J. Простагландин D2 модулирует кальциевые сигналы, индуцированные простагландином E2 в нейронах ганглиев задних корешков крыс. Neurosci Lett. 2015; 597: 159–63.
CAS PubMed Статья Google Scholar
Popp L, Haussler A, Olliges A, Nusing R, Narumiya S, Geisslinger G, Tegeder I. Сравнение ноцицептивного поведения у мышей с нокаутом простагландинов E, F, D, простациклина и тромбоксана.Eur J Pain. 2009; 13: 691–703.
CAS PubMed Статья Google Scholar
Гиббс Дж. Л., Диоген А., Харгривз К. М.. Нейропептид Y модулирует действие брадикинина и простагландина E2 на ноцицепторы тройничного нерва посредством активации рецепторов Y1 и Y2. Br J Pharmacol. 2007; 150: 72–9.
CAS PubMed Статья Google Scholar
Николсон Т.А., Фостер А.Ф., Беван С., Ричардс С.Д.Простагландин E2 сенсибилизирует первичные сенсорные нейроны к гистамину. Неврология. 2007; 150: 22–30.
CAS PubMed Статья Google Scholar
Тай Х. Х., Энсор С. М., Тонг М., Чжоу Х., Ян Ф. Ферменты, катаболизирующие простагландины. Простагландины Other Lipid Mediat. 2002; 68–69: 483–93.
CAS PubMed Статья Google Scholar
Чжан Ю., Десаи А., Ян С.Ю., Бэ КБ, Антчак М.И., Финк С.П., Тивари С., Уиллис Дж. Э., Уильямс Н.С., Доусон Д.М. и др.РЕГЕНЕРАЦИЯ ТКАНЕЙ. Ингибирование расщепляющего простагландин фермента 15-PGDH усиливает регенерацию тканей. Наука. 2015; 348: ааа2340.
PubMed PubMed Central Статья CAS Google Scholar
Антонова М., Винеке Т., Олесен Дж., Ашина М. Простагландины при мигрени: обновленная информация. Curr Opin Neurol. 2013; 26: 269–75.
CAS PubMed Статья Google Scholar
Чжан XC, Страссман AM, Бурштейн Р., Леви Д. Сенсибилизация и активация внутричерепных менингеальных ноцицепторов медиаторами тучных клеток. J Pharmacol Exp Ther. 2007. 322: 806–12.
CAS PubMed Статья Google Scholar
Harriott AM, Gold MS. Электрофизиологические свойства афферентов твердой мозговой оболочки в отсутствие и в присутствии медиаторов воспаления. J Neurophysiol. 2009; 101: 3126–34.
PubMed PubMed Central Статья Google Scholar
Вон А.Х., Золотой MS. Ионные механизмы, лежащие в основе индуцированной медиатором воспаления сенсибилизации афферентов твердой мозговой оболочки.